
Prirodna baština
Projekt COAST
Očuvanje i održivo korištenje biološke i krajobrazne raznolikosti na dalmatinskoj obali putem održivog razvitka obalnog područja
Morska bioraznolikost otoka Biševa i jugoistočne strane otoka Visa
Stručna podloga za održivo upravljanje

Zadatak 1. Izrada stručne podloge nužne za upravljanje morskim područjima otoka Biševa i jugoistočne strane otoka Visa kao dijelovima Nacionalne ekološke mreže_
1. UVOD
Oceanografske značajke
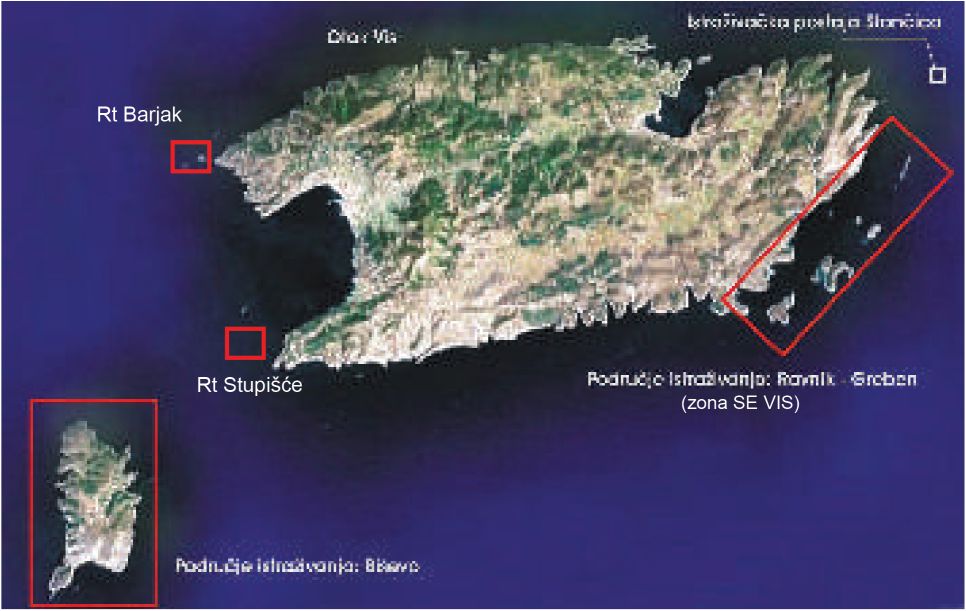
Otoci Biševo i Vis pripadaju skupim vanjskih otoka srednjeg Jadrana. Osnovni oceanografski podaci kojima se može opisati područje Biševa i Visa sakupljaju se već desetljećima od strane Instituta za oceanografiju i ribarstvo na postaji Stončica (Slika 1).
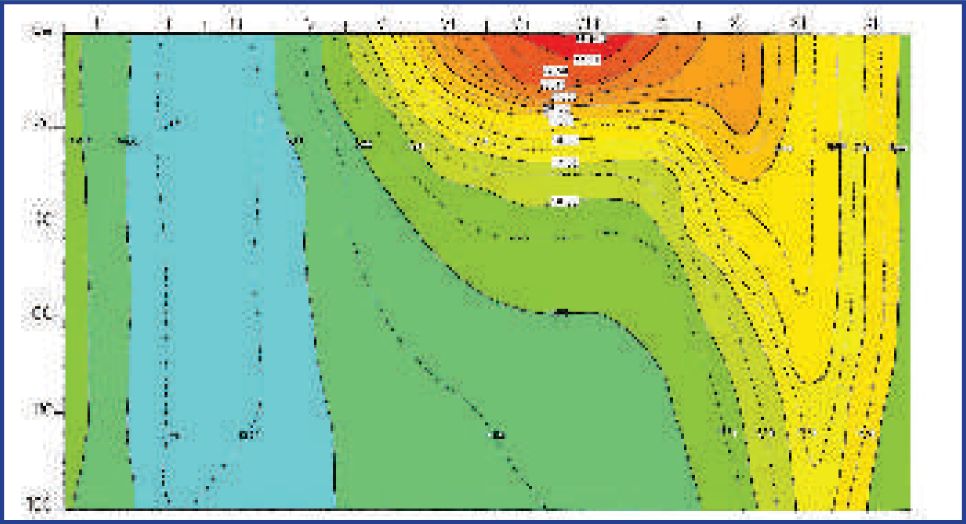
Temperatura mora ovog područja mijenja se po dubini tijekom godine što je vidljivo na grafu prosječnih temperatura za postaju Stončica (Slika 2).
U prosjeku od studenog do travnja, temperatura mora se po dubini ne mijenja te vlada izotermija. Početkom svibnja razvija se termoklina (naglo smanjenje temperature s povećanjem dubine), a koja je stabilna do rujna, kada se ponovo počinje razvijati izotermija. Potpuna izotermija od površine do dna se razvija na temperaturi od 17°C koncem studenog. Zimskim se hlađenjem do kraja ožujka temperatura čitavog stupca mora spusti još za oko 4°C.
Prema podacima o površinskim temperaturama na postaji Stončica od 2004. do kraja 2007. godine, maksimalne izmjerene vrijednosti bile su oko 25,5°C, a najniže oko 13,1°C (Slika 3).
Površinska strujanja mijenjaju se također sezonski, a njima pomažu i sezonske promjene režima vjetra. Zimi prevladava sjeverozapadni smjer, u proljeće sjeverni, ljeti jugoistočni, a u jesen jugozapadni smjer površinskih struja (Slika 4). Za vrijeme zimskog strujanja zapadnog smjera, a u vrijeme najjačeg priljeva rijeke Neretve, utjecaj rijeke se opaža i kod otoka Visa.
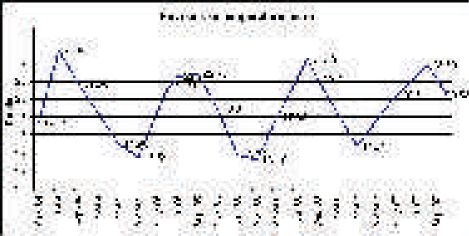
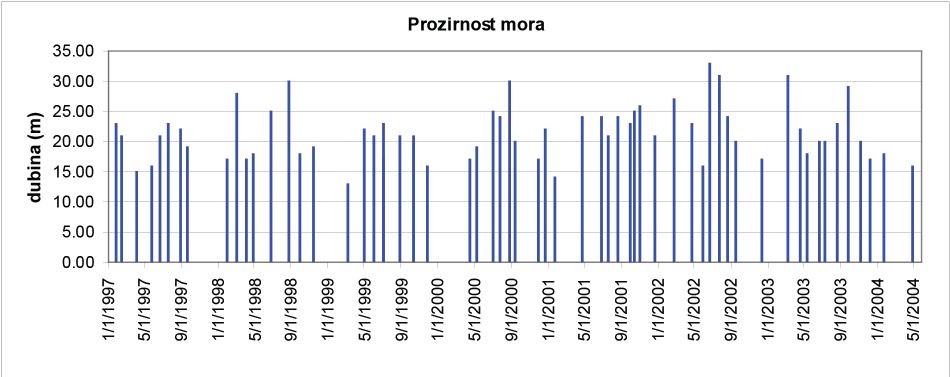
Na ovom području je prozirnost mora velika, a boja plava u raznim tonovima. Plava boja indicira visoki salinitet i malu količinu raspršenih čestica. Vrijednosti prozirnosti mora na istraživačkoj postaji Stončica od 1997. do 2005. kolebala su od minimalnih 13 do maksimalnih 33 metra dubine. Prosječne vrijednosti su oko 21,5 metara (Slika 5). Za usporedbu, prozirnosti mora na srednjojadranskom pragu su od minimalnih 14 do maksimalnih 50 metara dubine, a prosječna prozirnost je 31,1 m. Maksimalne su vrijednosti izmjerene u IX, II i III mjesecu, a minimalne u V i VIII. Na otvorenom sjevernom Jadranu prozirnost je od 5 do 39 m, a na području južnog dubokog Jadrana od 17 do 56 m.
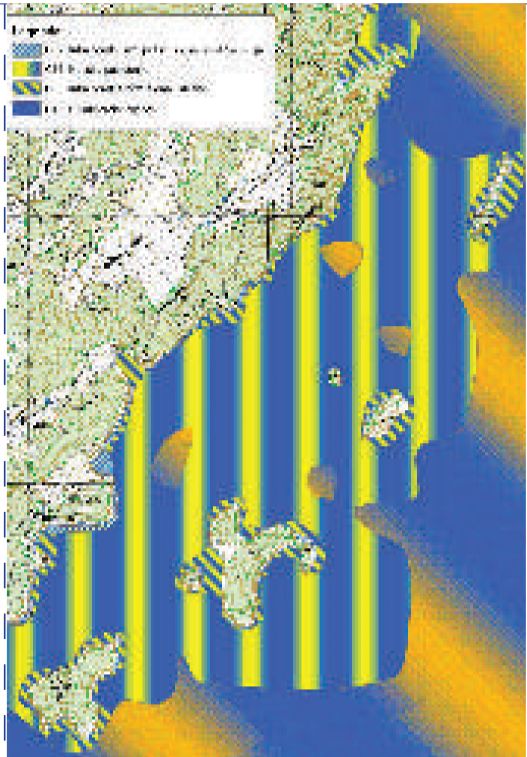
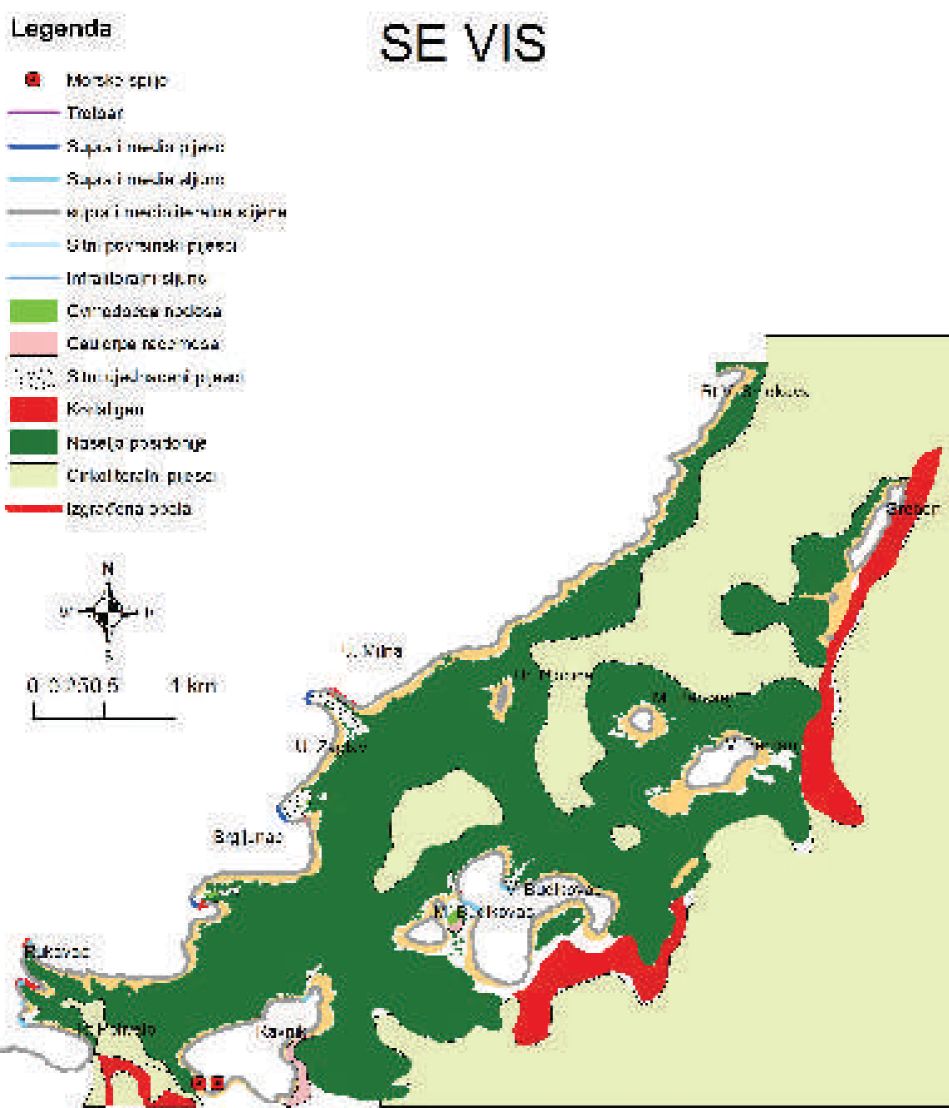
Slika 6. Karta morskih staništa istočne strane otoka Visa od otočića Ravnika do Grebena.
Biološke značajke
Ne postoje objavljeni znanstveni radovi u kojim se ciljano bavilo inventarizacijom vrsta i staništa morskog područja Biševa i predjela od Grebena do Ravnika. U posljednje je vrijeme područje oko Grebena obrađivano u sklopu nekoliko diplomskih radova s tematikom biometrije i ekologije vrste Posidonia oceanica.
Pojedini znanstveni radovi samo se dotiču ovog područja na način da tamo bilježe pojedine vrste, ali u sklopu geografsko širih istraživanja.
Ovdje obuhvaćamo takve podatake, ali i naše neobjavljene podatke sakupljene tijekom prijašnjih istraživanja ovog područja te dajemo pretpostavke za postojanje određenih vrsta i stanišnih tipova na istraživanom području.
Morska staništa
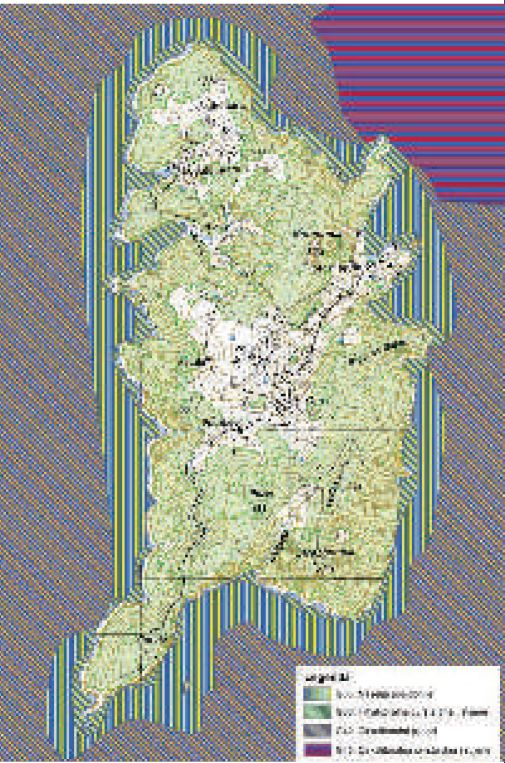
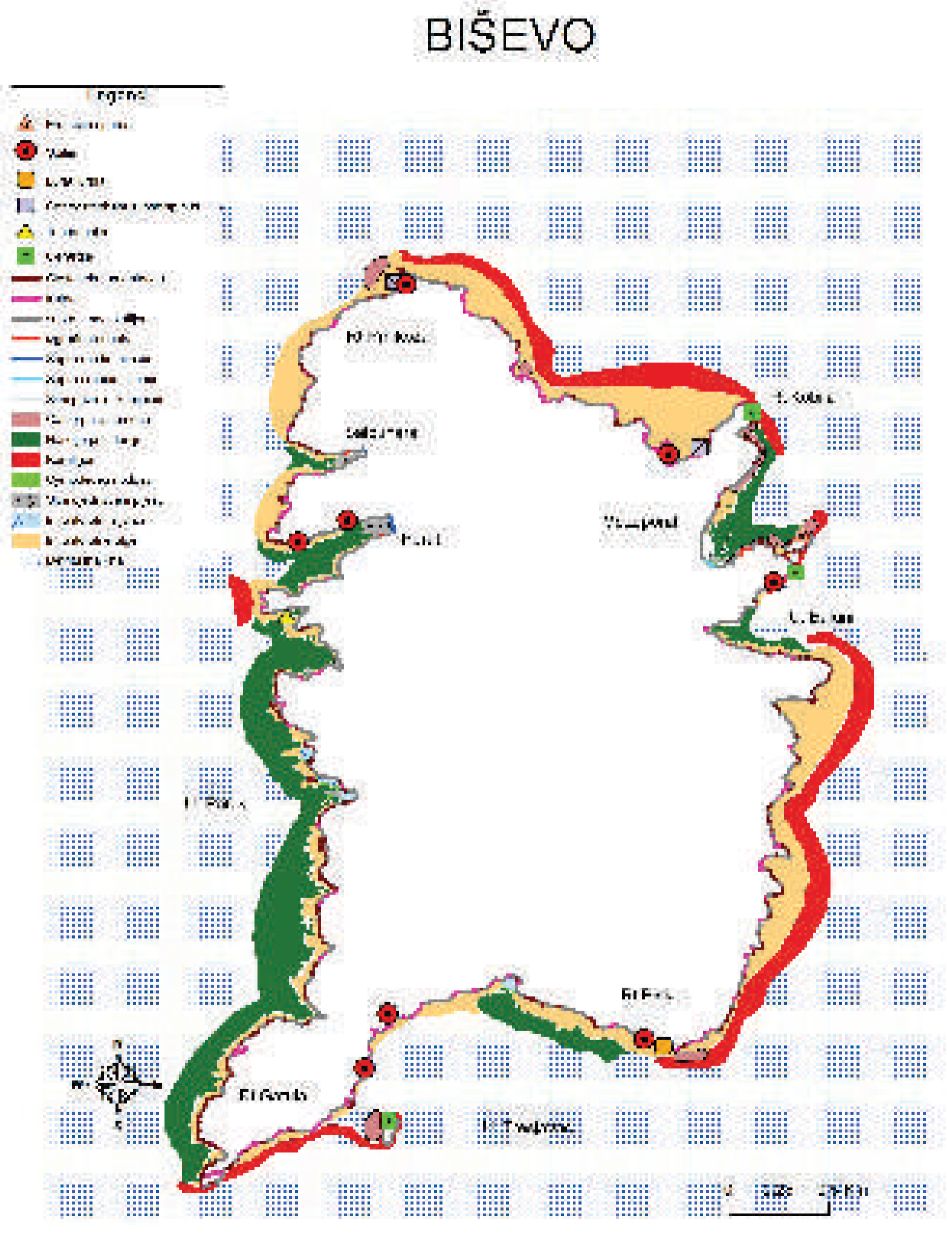
Slika 7. Karta morskih staništa otoka Biševa.
Grebena obrađivano u sklopu nekoliko diplomskih radova s tematikom biometrije i ekologije vrste Posidonia oceanica.
Pojedini znanstveni radovi samo se dotiču ovog područja na način da tamo bilježe neke ciljane vrste, ali u sklopu geografsko širih istraživanja.
Prema karti staništa DZZP, morska staništa prisutna na području od Ravnika do Grebena su:
G.3.2. Infralitoralni sitni pijesci s više ili manje mulja
G.3.5. Naselja posidonije
G.3.6. Infralitoralna čvrsta dna i stijene
G.4.2. Cirkalitoralni pijesci
Na području Biševa prema karti staništa DZZP,
prisutna su morska staništa:
G.3.5. Naselja posidonije
G.3.6. Infralitoralna čvrsta dna i stijene
G.4.2. Cirkalitoralni pijesci
Ne postoje objavljeni znanstveni radovi u kojim se ciljano bavilo inventarizacijom vrsta i staništa morskog područja Biševa i predjela od Grebena do Ravnika. U posljednje je vrijeme područje oko
Naselja morske cvjetnice Posidonia oceanica široko su rasprostranjena oko otoka Biševa i na istočnoj strani otoka Visa, posebno na području od otočića Ravnika do Grebena (Slike 6 i 7).
Prikaz morskih staništa na ovoj je karti indikativan, a dobiven je metodom prostornog modeliranja. Prema online Bazi podataka Nacionalna ekološka mreža za istraživana područja postoje sljedeći podaci:
| Šifra i naziv područja: HR3000096 # , JI strana o. Visa | ||
| Ciljevi očuvanja | ||
| Stanišm tipovi | ||
| NKS šifra | NATURA šifra | stanišni tip |
| G.3.5. | 1120* | Naselja posidonije |
| Mjere zaštite | ||
| 25: 31: 32: 132: 133: 135_ |
| Šifra i naziv područja: HR3000098 # , Biševo more | ||
| Ciljevi očuvanja | ||
| Stanišni tipovi | ||
| NKS šifra | NATURA šifra | stanišni tip |
| G.3.5. | 1120* | Naselja posidonije |
| um | ΊΠΓ70 | Grebeni |
| 1J33Ü | Morske špilje | |
| F.3. | Šljunkovita morska obala | |
| 1110 | Pješčana dna | |
| 1140 | Muljevite i pješčane plićine | |
| F.2. | Pjeskovita morska obala | |
| Mjere zaštite11 ; 21 ; 22; 23; 25; 31 ; 32; 132; 133; Ostalo: očuvati povoljne stanišne uvjete_ |
2. METODOLOGIJA
Metodologija koja se koristila za inventarizaciju morske biološke raznolikosti uključivala je sakupljanje, analizu i interpretaciju postojećih te sakupljanje, analizu i interpretaciju novih podataka tijekom provedbe projekta te njihovo zajedničko uključivanje u izvještaj.
Kako bi se donijele preporuke za održivi razvoj Viškog područja, obavljena je radionica s lokalnom upravom i interesnim skupinama (ribari, turistički djelatnici i dr.) na kojoj je razmatran niz prijedloga.
Popularna interpretacija (izrada filmova, brošura i info-ploča) obavljena je u suradnji s profesionalnim grafičarima.
Terensko istraživanje
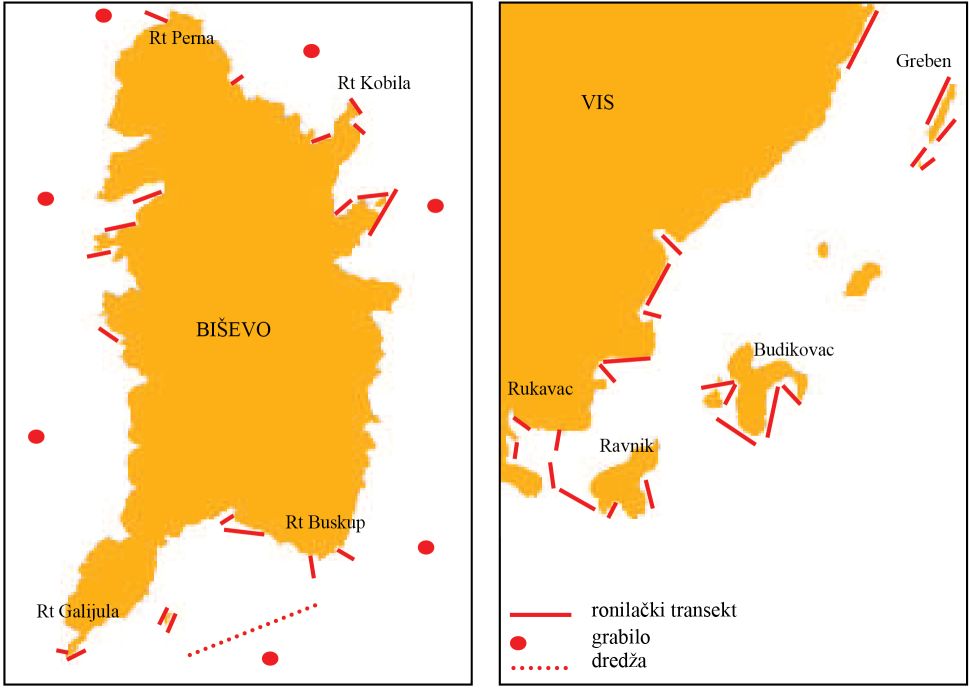
Terensko je istraživanje obavljeno na dva područja i to 1) Biševo i 2) jugoistočna strana otoka Visa od otočića Greben do otočića Ravnik (dalje u teksu: zona SE) (Slika 1). Terenski rad se obavljao sezonski uz korištenje istraživačkog broda BIOS, brodice tipa Calafuria i gumenog čamca (Slika 8, 9).
Terenski je rad uključivao:
identifikaciju morskih staništa sukladno Nacionalnoj klasifikaciji staništa i EU direktivi o staništima
– kartiranje staništa i najvažnijih (zaštićenih, ugroženih, karakterističnih, alohtonih, invazivnih, vrijednih) morskih vrsta. Za određivanje pozicije koristio se uređaj Vectorsensor CSI Wireless, a dubine WHS600-I-UG62 ADCP RD Instruments
– identifikaciju mogućih opasnosti za bioraznolikost istraživanog područja

Slika 8. Terensko istraživanje. Kartiranje trotoara i alge Cvstoseira amantacea var. spicata na Biševu uz pomoć gumenjaka (a), analiza lovine poponica u Rukavcu (b), kartiranje posidonije uz pomoć brodice Navicula s istočne strane Budikovca (c), istraživački brod Bios na južnom dijelu Biševa (d), uzorko-vanje grabilom iz broda Bios na južnom dijelu Biševa (e) i ispiranje materijala sakupljenog dredžom (f), ronilački transekt uz hrid Totac (g), obrada uzoraka u brodu Bios (i), snimanje i uzorkovanje u korali-genu (j, h).
Slika 9. Područje istraživanja: Biševo i jugoistočna strana otoka Visa od otočića Greben do otočića Ravnik (dalje u teksu: zona SE). Označeni su ronilački transekti i mjesta uzorkovanja dredžom i grabilom.
– identifikaciju hot spot područja biološke raznolikosti
– sakupljanje foto i video materijala korištenjem digitanih fotoaparata i kamera mini DV formata
– konzultaciju s lokalnom upravom i interesnim skupinama (ribari, ronilački centri, turistički djelatnici i dr.).
Skupljanje biološkog materijala obavljalo se:
– pomoću autonomnih ronioca do dubina od 40 m (bentoski organizmi, identifikacija i kartiranje zajednica i staništa, vizualni cenzus riba, fotografiranje i snimanje)
– pomoću dredže i grabila za dubine od 30 do 90 m (bentoski organizmi)
Sve potrebne dozvole za ronjenje i sakupljanje organizama bile su nabavljene prije terenskog rada.
Laboratorijski i uredski rad
2. obrada sakupljenog biološkog materijala
3. analiza podataka
4. izrada GIS karata staništa i odabranih svojti
5. izrada inventarnih lista zabilježenih organizama iz skupina:
a. fitobentos
b. zoobentos
c. fitoplankton
d. zooplankton
e. morski sisavci
6. odabir i obrada foto materijala
7. odabir i montiranje video materijala
8. izrada edukacijskih brošura za tri odabrana lokaliteta
9. izradu info ploče o podmorju Biševa
10. formuliranje preporuka za očuvanje, održivo korištenje i upravljanje istraživanim područjem
11. formuliranje preporuka za daljnje istraživanja i aktivnosti uz okvirni troškovnik
12. formuliranje preporuka za aktivnosti praćenju istraživanog područja uz okvirni troškovnik.
Laboratorijski i uredski rad uključivao je:
1. sakupljanje objavljenih podataka (znanstveni radovi, izvještaji, i dr.)
Izrada karte staništa i pojedinih vrsta
Karte rasprostranjnosti napravljene su u GIS formatu za prevladavajuća staništa i neke rijetke i zaštićene svojte (Tablica 1).
Rasprostranjenost naselja posidonije određena je uz pomoć zračnih snimaka (orto foto) i dubinskih transekata na odabranim lokalitetima. Gornji (plići) rub livade određen je iz orto foto snimaka. Rub je ručno ucrtan uz pomoć tableta, a linija digitalizirana i georeferencirana (Slika 10). Donji rub (dublji) je određen na osnovi in situ određene maksimalne dubine (32 – 34 m ) na nekoliko odabranih transekata, a u kartu ucrtan na izobati od 30 m, odnosno na dubini od koje preostaju samo pojedinačni čuperci.
Pojedina područja s dubljim gornjim rubom posidonije koji nisu vidljivi iz zračnih snimaka su kartirana pomoću autonomnih ronioca. Ronioc je uz pomoću ronilice slijedio rub posidonije i za sobom povlačio plutaču. Plutaču je slijedila brodica opremljena GPS sučeljem (Slika 11). Tako ucrtana putanja pridodana je GIS sloju naselja posidonije. Ova je metoda nepreciznija od ucrtavanja ruba na osnovi zračnih snimaka i primjenjivala se samo iznimno.
Da bi se odredila rasprostranjenost trotoara i alge Cystoseìra amantacea var. spicata na Biševu, uz pomoć malog čamca je obilažena obala, a nalazi ucrtavani na aero snimke. Zabilježbe su naknadno digitalizirane i georefrencirane. Na isti je način obavljeno kartiranje stjenovite, pješčane, šljunčane i antropogene obale.

Infralìtoralni šljunci i pijesci kartirani su iz zračnih snimaka nakon provjere staništa na terenu. Rasprostranjenost koraligena određena je na osnovi terenskih zabilježbi i batimetrije. NA područjima koja nisu pregledana autonomnim roniocima, pretpostavka je da se da je koraligen razvijen ako je dubina veća od 30 m i ako se dno strmo obrušava, a što onemogućava razvoj sedimentnih dna. Sva dublja dna od 30 m s blagim nagibom pripadaju sedimentnim dnima cirkalitorala. Postojanje ovih dna potvrđeno je uzorkovanjem dredžom i grabilom.
Nalazi alge Caulerpa racemosa var. cylindracea tijekom ronjenja ucrtani su u GIS sloj. Za pretpostaviti je da je njena rasprostranjenost u prirodi daleko veća nego je prikazana na kartama rasprostranjenosti.

Tablica 1. Popis staništa i svojti za koje su izrađene G1S karte rasprostranjenosti.
| Rukavac | Biševo |
| Supralitoralne i mediolitoralne stijene (F.4.2., G.2.4.)Supralitoralni i mediolitoralni pijesci (F.2.2., G.2.2.)Supralitoralni i mediolitoralni šljunci (F.3.2., G.2.3.)Izgrađene obale (F.5.1.2.1.)Trotoar Morske špiljeSitni površinski pijesci (G.3.2.1.)Sitni ujednačeni pijesci (G.3.2.2.)Cymodocea nodosa (G.3.2.2.1.)Infralitoralni šljunci (G.3.4.1.)Naselja posidonije (G.3.5.)Infralitoralne alge (G.3.6.1.)Caulerpa racemosa (G.3.8.6.2.)Cirkalitoralni pijesci (G.4.2.)Koraligen (G.4.3.1.) | Supralitoralne i mediolitoralne stijene (F.4.2., G.2.4.)Supralitoralni i mediolitoralni pijesci (F.2.2., G.2.2.)Supralitoralni i mediolitoralni šljunci (F.3.2., G.2.3.)Izgrađene obale (F.5.1.2.1.)Trotoar Morske špiljeSitni površinski pijesci (G.3.2.1.)Sitni ujednačeni pijesci (G.3.2.2.)Cymodocea nodosa (G.3.2.2.1.)Infralitoralni šljunci (G.3.4.1.)Naselja posidonije (G.3.5.)Infralitoralne alge (G.3.6.1.)Cystoseira amantacea var. spicata (G.3.6.1.2.) Caulerpa racemosa (G.3.8.6.2.)Obalna detritusna dna (G.4.2.2.)Koraligen (G.4.3.1.)Luria lurida Erosaria spurca Tonna galea Gerardia savaglia Centrostephanus longispinus |
Literatura:
Antolić B., A. Špan, AŽuljević, Vukovič A. 2001. Check list of the benthic marine macroalgae on the eastern Adriatic coast: I. Chlorophyta. Acta Adriat., 42 (2): 43-58.
Bakran-Petricioli T., 2007. Morska staništa. Državni zavod za zaštitu prirode.
Buljan M. i Zore-Armanda M., 1971. Osnovi oceanografije i pomorske meteorologije. Institut za oceanografiju i ribarstvo – Split: 424 str.
Ercegović A. (1952). Jadranske cistozire. Institut za oceanografiju i ribarstvo. Split. 212 str.
Ercegović A. (1980). Etude comparative de la végétation de basses eaux et de celle des eaux profondes de l’Adriatique. Acta Adriat., 21: 11-40. Gamulin-Brida H., Jardas I., Špan A. (1987). problemi endema mora s posebnim osvrtom na Jadran. Akademija nauka i umjetnosti Bosne i Hercegovine. Posebna izdanja. Odjeljenje prirodnih i matematičkih nauka. Knjiga LXXXIII. 57-73.
Gamulin-Brida H., Jardas I., Špan A., 1987.
Problemi endema mora s posebnim osvrtom na Jadran. Akademija nauka i umjetnosti Bosne i Hercegovine. Posebna izdanja. Odjeljenje prirodnih i matematičkih nauka. Knjiga LXXXIII. 57-73.
Peres J-M., Gamulin Brida H., 1973. Biološka oceanografija. Školska knjiga, Zagreb: 493 str.
Regner, D. (1981) The changes in seasonal oscillations of copepods in the central Adriatic. Rapp. Comm. hit. Mer Médit., 27 (7): 177-179.
Regner, D. (1985) Seasonal and multiannual dynamics of copepods in the middle Adriatic. Acta Adriat., 26 (2): 11-99.
Regner, D. (1991) Long-term investigations of copepods (zooplankton) in the coastal waters of the easterm Middle Adriatic. Acta Adriat. 32 (2): 631-740
Šolić M., Krstulović N., Marasović I., Baranović A., Pucher-Petković T., Vučetić T. (1997) Analysis of time series of planktonic communities in the Adriatic Sea: distinguishing between natural and man-induced changes. Oceanologica Acta 20 (1): 131-143.
3. REZULTATI
Popis staništa istraživanog područja
Provedenim istraživanjima tijekom projekta COAST utvrđeno je na istraživanom području ukupno 23 biocenoze i zajednice i 13 nižih kategorija (facijesi, asocijacije i dr.) (Slika 12a, b). Pregled ovih staništa, njihova rasprostranjenost i značajke navode se u narednim poglavljima.
F. Morska obala
F.2. Pjeskovita morska obala
F.2.2. Supralitoralni pijesci
E2.2.1. Biocenoza supralitoralnih pijesaka
F.3. Sljunkovita morska obala F.3.2. Supralitoralni šljunci i kamenje F.3.2.1. Biocenoza sporosušećih nakupina ostataka morske vegetacije.
F.4. Stjenovita morska obala
F.4.2. Supralitoralne stijene
E4.2.1. Biocenoza supralitoralnih stijena
F.5. Antropogena staništa morske obale F.5.1. Antropogena staništa morske obale F.5.1.1. Zajednice morske obale na pomičnoj podlozi pod utjecajem čovjeka F.5.1.2. Zajednice morske obale na čvrstoj podlozi pod utjecajem čovjeka
F. 5.1.2.1. Izgrađene i konstruirane obale
G. More
G.2. Mediolitoral
G.2.2. Mediolitoralni pijesci
G.2.2.1. Biocenoza mediolitoralnih pijesaka
G.2.3. Mediolitoralni šljunci i kamenje
G.2.3.1. Biocenoza mediolitoralnih dna s
krupnim detritusom
G.2.3.1.1. Facijes s naslagama mrtvog lišća vrste Posidonia oceanica i drugih morskih cvjetnica G.2.4. Mediolitoralno čvrsto dno i stijene G.2.4.1. Biocenoza gornjih stijena mediolitorala
G.2.4.1.1. Asocijacija s vrstom Bangia atropurpurea
G.2.4.2. Biocenoza donjih stijena mediolitorala
* asocijacije s inkrustriranim crvenim algama nije moguće specificirati
G.2.4.3. Biocenoza mediolitoralnih špilja
G.2.5. Antropogena staništa u mediolitoralu G.2.5.1. Zajednice mediolitorala na pomičnoj podlozi pod utjecajem čovjeka
G.2.5.2. Zajednice mediolitorala na čvrstoj podlozi pod utjecajem čovjeka
G.2.5.2.1. Facijesi mediolitorala betoniranih i izgrađenih obala (luke, lučice, brodogradilišta) i ostalih ljudskih konstrukcija u moru
G.3. Infralitomi
G.3.2. Infralitoralni sitni pijesci s više ili manje mulja
G.3.2.1. Biocenoza sitnih površinskih pijesaka G.3.2.2. Biocenoza sitnih ujednačenih pijesaka
G.3.2.2.1. Asocijacija s vrstom Cymodocea nodosa
G.3.3. Infralitoralni krupni pijesci s više ili manje mulja
G.3.4. Infralitoralno kamenje i šljunci G.3.4.1. Biocenoza infralitoralnih šljunaka
G.3.5. Naselja posidonije
G.3.5.1. Biocenoza naselja vrste Posidonia oceanica
G.3.5.1.3. Facijes mrtvih naslaga rizoma posidonije bez epiflore
G.3.6. Infralitoralna čvrsta dna i stijene G.3.6.1. Biocenoza infralitoralnih alga
G.3.6.1.1. Degradirani facijes s inkrustrirajućim algama i ježincima
G.3.6.1.2. Asocijacija s vrstom Cystoseira
amantacea var. spicata
G.3.8. Antropogena staništa u infralitomi!!
G.3.8.6. Infralitoralne zajednice s invazivnim vrstama
G.3.8.6.2. Zajednice s vrstom Caulerpa racemosa
G.4. Cirkalitoral
G.4.2. Cirkalitoralni pijesci
G.4.2.2. Biocenoza obalnih detritusnih dna
G.4.3. Cirkalitoralna čvrsta dna i stijene G.4.3.1. Koraligenska biocenoza G.4.3.1.5. Facijes s vrstom Eunicella cavolinii G.4.3.1.6. Facijes s vrstom Eunicella singularis G.4.3.1.8. Facijes s vrstom Paramuricea clavata G.4.3.1.9. Facijes s vrstom Parazoanthus axinellae G.4.3.2. Biocenoza polutamnih špilja G.4.5. Antropogena staništa u cirkalitoralu G.4.5.2. Podmorska arheološka nalazišta
G.5.3.2. Biocenoza špilja i prolaza u potpunoj tami
Analiza zabilježenih staništa istraživanog područja

F. Morska obala
Većina obale je stjenovita (F.4. Stjenovita morska obala), dok manji dio zauzima pjeskovita, šljunkovita i antropogena obala (F.2. Pjeskovita morska obala, F.3. Šljunkovita morska obala,
F.5. Antropogena staništa morske obale).
F.4. Stjenovita morska obala
F.4.2. Supralitoralne stijene
F.4.2.1. Biocenoza supralitoralnih stijena
Ova je biocenoza iznimno dobro razvijena zbog izloženosti obale valovima. Tako npr. s istočne
i južne strane Biševa pojas supralitorala može iznositi i pet metara u visinu (Slika 12).
Analiza supralitoralnih modrozelenih alga, a koje daju izgledu obale specifičnu sivu obojenost, nije rađena jer za ovu skupinu organizama u Hrvatskoj ne postoje eksperti.
Od životinja koje žive na ovom području tipični organizmi su: puževi Melarhaphe neritoides, izopodni račić Ligia italica i ciripedni račića Chthamalus depressus (Slika 13).
Biocenoza na istraživanom području nije ugrožena.
Slika 12. Stjenovita morska obala. Sjeverna strana otočića Greben i jugoistočna strana otoka Biševo. Tamnosivo područje označava zonu supralitorala koja dostiže i nekoliko metara visine zbog izloženosti valovima.
Slika 13. Tipični organizmi supralitorala: puževi Melarhaphe neritoides, izopodni račić Ligia italica i ciripedni račić Chthamalus depressus.
Biocenoze supralitoralnih i mediolitoralnih pijesaka
F. 2.2.1. Biocenoza supralitoralnih pijesaka
G. 2.2.1. Biocenoza mediolitoralnih pijesaka
Ove su biocenoze razvijene u uvalama Porat i Salbunara na Biševu te u uvalama Zaglav i Milna u SE zoni (Slika 14). Između ove dvije biocenoze koje se pružaju jedna iznad druge, teško je odrediti graničnu liniju.
Detaljna faunistička analiza ovih biocenoza nije provedena premda j e unutar naslaga lišća posidonije uočen amfipodni račić Talitrus saltator. Tijekom jeseni na ovo stanište more nanese znatne količine ostataka listova morske cvjetnice posidonije. Ove naplavine imaju bitnu ekološku funkciju, jer pridonose donosu organske tvari, ali imaju i zaštitnu funkciju u sprječavanju erozije obale uslijed valovanj a. Snažni valovi uslij ed j užnih vj etrova koj i se razvijaju tijekom jeseni mogu značajno djelovati na eroziju ovakvih staništa, a listovi posidonije imaju zaštitnu funkciju (Slika 15).




Ova se staništa koriste kao plaže, mjesta za izvlačenje i suhi vez brodica. Nj ihovo iskorištavan]’ e za plaže je intenzivno tijekom ljetnog razdoblja, te su ove zajednice pod velikim pritiskom ljudskih aktivnosti.
Naplavine smeća tijekom jeseni i zime znaju biti znatne, ali se uklanjaju prije turističke sezone i uglavnom nemaju negativan utjecaj na zajednicu. Korištenje ovih staništa za plaže je realnost. Međutim, potrebno je barem spriječiti uklanjanja naplavljenih listova posidonije tijekom jeseni i zime.
Potrebno j e paziti da razni zahvati u prirodi ne utj eču negativno na ova staništa kao što je trenutno slučaj s izradom potpornog zida u uvali Milna pri čemu je zemljom i kamenjem nasuta gotovo polovica plaže (Slika 16).
Biocenoze supralitoralnih i mediolitoralnih šljunaka
F.3.2.1. Biocenoza sporosušećih nakupina ostataka morske vegetacije
U SE zoni Visa ova staništa su razvijena u uvali Rukavac, na južnoj i sjevernoj strani otočića Budikovca, a desetak metara dugačka šljunčana obala razvijena je i na sjevernom rtu otočića Ravni ka (Slika 17).
Na otoku Biševo ova su staništa dobro razvijena u uvali Mezuporat, a manja, desetak metara dugačka šljunčana obala razvijene je u uvali Potok na zapadnoj strani otoka (Slika 18 i 19).
Između ove dvije biocenoze koje se pružaju jedna iznad druge, teško je odrediti graničnu liniju. Tijekom jeseni i zime ovdje more nanosi značajne količine listova posidonije pa je u supralitoralu razvijena Biocenoza sporosušećih nakupina ostataka morske vegetacije (F.3.2.1.).

Ove nakupine listova prisutne su tijekom cijele godine za razliku od mediolitorala gdje se značajno nakupljanje detritusa odvija samo tijekom jeseni i zime (G.2.3.1. Biocenoza mediolitoralnih dna s krupnim detritusom) kada je dobro razvijen facijes s naslagama mrtvog lišća vrste Posidonia oceanica i drugih morskih cvjetnica (G.2.3.1.1.) (Slika 21).
Isto kao i pješčana obala, šljunčana se obala koristi kao plaža, mjesto za izvlačenje i suhi vez brodica (Slika 21). Njihovo iskorištavanje za plaže je intenzivno tijekom ljetnog razdoblja kada su ove zajednice pod velikim pritiskom ljudskih aktivnosti.
Naplavine smeća tijekom jeseni i zime znaju biti znatne, ali se uklanjaju prije turističke sezone i uglavnom nemaju negativni utjecaj na zajednicu (Slika 20).
Korištenje ovih staništa za plaže je realnost. Međutim, potrebno je barem spriječiti uklanjanja naplavljenih listova posidonije tijekom jeseni i zime.
Potrebno je paziti da razni zahvati u prirodi ne utječu negativno na ova staništa kao što je izgradnja mula u uvali Rukavac. Antiobraštajne boje koje se uklanjanju s brodova u suhom vezu mogu negativno djelovati na osjetljiv ekosustav šljunčanih zajednica.




F.5. Antropogena staništa morske obale
F.5.1. Antropogena staništa morske obale
F.5.1.1. Zajednice morske obale na pomičnoj
podlozi pod utjecajem čovjeka
F.5.1.2. Zajednice morske obale na čvrstoj
podlozi pod utjecajem čovjeka
F.5.1.2.1. Izgrađene i konstruirane obale
G.2.4. Mediolitoralno čvrsto dno i stijene
G.2.4.1. Biocenoza gornjih stijena mediolitorala
O va j e biocenoza pod utj ecaj em moraj edino tij ekom zalijevanja valova. Zbog toga je izložena znatnom sušenju i ekstremnim ekološkim prilikama zbog kojih ovdje uspijeva mali broj organizama. Zonom dominiraju endolitske cijanobakterije koje stijeni daju maslinastosmeđu obojenost. Od životinja tu su priljepci roda Patella spp., ciripedni račići Chthamalus stellatus i C. montagui. U procjepima se razvija crvena alga Catenella caespitosa, a na stijenama u uvali Rukavac tijekom proljeća se razvija crvena alga Bangia atropurpurea, a time i asocijacija s vrstom Bangia atropurpurea (G.2.4.1.1.) (Slika 24) koja indicira organsko onečišćenje najvjerojatnije od kanalizacijskih ispusta ili septičkih jama.

Pješčane i šljunčane obala su pod utjecajem čovjeka. Koriste se za izvlačenje i suhi vez brodica te kao plaže. Njihovo korištenje je intenzivno tijekom ljeta kada su staništa pod velik pritiskom, dok se tijekom jeseni, zime i proljeća biocenoze obnavljaju. Izgrađene obale prisutne su u gotovo svim uvalama, ali je njihov udio za sada minimalan, a u funkciji su priveza brodova (Slike 22 i 23).



G.2.4.2. Biocenoza donjih stijena mediolitorala
Donji mediolitoral pod utjecajem je mora izmjenom plime i oseke te uslijed valovanja. Na područjima Biševa, u ovoj je biocenozi iznimno dobro razvijena biogena formacija „trotoar“ koju grade razne vrste inkrustriranih crvenih alga (Slika 25).
Prema Nacionalnoj klasifikaciji staništa nije moguće odrediti asocijacije crvenih alga u donjem mediolitoralu jer se iz službenih naziva NKS ne razumije o kojim je vrstama riječ. Naime, sistematika se često mijenjala, te je naziv za jednu vrstu ujedno i sinonim za drugu. Kako u nazivima asocijacija u NKS pri spomenu vrste nema i autora, nije moguće reći da li se radi o trenutno važećoj vrsti ili o sinonimu. Tako je npr. naziv vrste Lithophylhtm lichenoides Philippi 1837, sinonim za vrstu Lithophyllm byssoides (Lamarck) Foslie. Ujedno prema NKS postoje asocijacie s
Lithophyllum lichenoides i Lithophyllm byssoides pa je jasno kako dolazi do problema identifikacije. Trotoar je posebno razvijen u prolazu koji čini rt Gatula. Ovo je možda i najbolje razvijen trotoar u Jadranu pa zaslužuje posebnu pažnju i zaštitu. Trotoari se smatraju indikatorom čistog mora, a kako polako rastu, smatraju se ugroženim staništima.
Sve formacije trotoara detaljno su ucrtane u kartu staništa.
U području donjih stijena mediolitorala česte životinjske vrste su priljepci (Patella spp.), crvena moruzgva (Actinia equina), ogre (Osilinus turbinatus), dekapodni rakovi Eriphia verrucosa i Pachygrapsus marmoratus te riba babica Cotyphoblennius galerita (Slika 26).

Morske špilje
G.2.4.3. Biocenoza mediolitoralnih špilja G.4.3.2. Biocenoza polutamnih špilja G.5.3.2. Biocenoza špilja i prolaza u potpunoj tami
Morske su špilje jedna od karakteristika istraživanog područj a. N eke od nj ih su turistička atrakcij a kao što su Zelena špilja na Ravniku te Modra i Medvidova špilja na Biševu koje su ujedno i zaštićene u kategoriji spomenika prirode.
Tijekom kartiranja obale ucrtan je položaj manjih mediolitoralnih špilja na otoku Biševu.
U uvjetima smanjene svjetlosti razvija se manji broj svojti alga među kojima se u mediolitoralu ističu Hildenbrandia rubra. Catenella caespitosa i Phymatolithon lenormandii (Slika 27).
U infralitoralnim špiljama na područjima s dovoljno svijetla razvijaju se alge roda Pevssonnelia i zelena alga Palmophvllum crassum.
Ovdje dominira životinjska komponenta i to prvenstveno spužve, žamjaci i mahovnjaci (Slika 28). U dubljim dijelovima špilja gdje nema svjetla, alge se ne razvijaju, a prevladavaju spužve, sesilni mnogočetinaši, mahovnjaci i rakovi (Slika 29). Ova staništa imaju iznimni potencijal korištenja u turističke svrhe, međutim, pretjerane posjete mogu dovesti do narušavanja ovih krhkih ekosustava (BOX 1: Morske špilje).

Modra špilja na otoku Biševu posebna je po obojenosti koju stvara upadno svjetlo koje prolazi kroz podvodni otvor.



Z’ Modra špilja
Modra špilja nalazi se u maloj uvali Balun na istočnoj strani otoka Biševa. Spilja ima dva otvora, jedan manji, umjetno produbljen, kroz koji može proći čamac bez vesala i taj otvor nema utjecaja na rasvjetu u špilji. Drugi je otvor na južnoj strani špilje, oblika je svoda i mnogo širi te je ispod razine mora. Kroz taj podvodni otvor prodire u špilju sunčeva svjetlost koja se prolazeći kroz vodu najprije lomi, a onda se od dna špilje odbija. Kako se od dna reflektiraju pretežno modre zrake svjetlosti, predmeti su u špilji obojeni modrom, a predmeti u vodi imaju srebrnastu boju. Ova pojava nastaje u špilji samo onda kada sunčeve zrake padaju u smjeru podvodnog otvora odnosno između 11 i 12 sati.
V_J
Medvidina špilja
Medvidina špilj a j e građena od prostranog vanj skog dijela koji ribarima služi kao zaklon od bure i unutrašnjeg užeg i tamnijeg dijela. Ime je dobila po tuljanu, sredozemnoj medvjedici (Monachus albiventer) koji je u toj špilji znao boravit, a kojeg na Visu zovu morski covik.
Zelena špilja
Zelena špilja nalazi se na otočiću Ravniku. Svjetlo u špilju prodire kroz dva velika ulazna otvora, ali i kroz mali prolaz na vrhu špilje. Upravo taj otvor propušta usku zraku svjetla koja u špilji razvija posebne svjetlosne efekte. Zelena špilja na otočiću Ravniku, često je turističko odredište.


Zakonom zaštićeni organizmi zabilježeni u ovim biocenozama:
1. puž zupka Luria lurida
2. spužva špiljska sumporača Aplysina cavernicola
G.3. Infralitoral
Područje infralitorala obuhvaća morsko dno od granice normalnih oseka do najvećih dubina na kojima su razvijene morske cvjetnice i fotofilne alge. Na vanjskim otocima srednjeg jadrana donja granica infralitorala doseže dubine od oko 40 metara. Na istraživanom područje ona je na osvijetljenim područjima dosezala između 30 i 35 m dubine.
Sitni površinski i ujednačeni pijesci
G.3.2.1. Biocenoza sitnih površinskih pijesaka G.3.2.2. Biocenoza sitnih ujednačenih pijesaka
G.3.2.2.1. Asocijacija s vrstom Cymodocea nodosa
Ove se biocenoze prvenstveno nastavljaju na pješčanu obalu. Pojavljuju se u uvalama Porat i Saplunara na Biševu te u uvalama Milna i Zaglav u zoni SE. Biocenoza sitnih ujednačenih pijesaka javlja se kao manja površina unutar naselja posidonije, npr. između otočića Budikovca i Rukavca te u uvalama Mezuporat i Potok na Biševu. U biocenozi sitnih ujednačenih pijesaka povremeno se javlja i morska cvjetnica Cymodocea nodosa odnosno asocijacija s vrstom Cymodocea nodosa (G.3.2.2.1.) (Slika 30).
Ugroženost biocenoza sitnih površinskih pijesaka istraživanog područja ista je kao i biocenoze supralitoralnih i mediolitoralnih pijesaka. Tijekom ljetne sezone služe kao plaže pa su pod jakim ljudskim utjecajem.
Biocenoza sitnih ujednačenih pijesaka na istraživanom području nije ugrožena zbog ljudskih djelatnosti. Međutim, na nekim je mjestima zahvaćena invazivnom vrstom Caulerpa racemosa var. cylìndracea (južna strana otočića Budikovca, uvala Mezuporat) (Slika 31).
Zakonom zaštićeni organizmi zabilježeni u ovim biocenozama:
1. morska cvjetnica čvorasta morska resa
Cymodocea nodosa
2. školjkaš plemenita periska Pinna nobilis

Slika 31. Naselje vrste Cymodocea nodosa zahvaćeno invazivnom algom Caulerpa racemosa var. cvlindracea uz južnu stranu otočića Budikovca.
G.3.4.1. Biocenoza infralitoralnih šljunaka
Ova se biocenoza obično nastavlja na šljunčane plaže. Biocenoza je zbog karaktera naše obale općenito rijetka. Na istraživanom je području zabilježena uz otočić Budikovac, te na Biševu u uvali Potok (Slika 32). Zbog nestabilnih ekoloških uvjeta uslijed valjanja oblutaka, ova staništa naseljava relativno mali broj specijaliziranih organizama. Neke inkrustrirane alge naseljavaju donje strane oblutaka. Na istraživanom području nije direktno ugrožena.

G.3.5. Naselja posidonije
G.3.5.1. Biocenoza naselja vrste Posidonia oceanica
G.3.5.1.3. Facijes mrtvih naslaga rizoma posidonije bez epiflore
Naselja posidonije dobro su razvijena na SE zoni (BOX 2). Na području Biševa razvijena su u uvali Mezuporat, u uvali Baiunte na većem dijelu zapadne strane otoka (Slika 33). Naselja se razvijaju gotovo od površine, kao u uvali Mezuporat, pa do 32 m dubine. U nekim uvalama naselje nije kompaktno već se razij a u obliku manj ih krpastih kolonij a (Slika 35). Na pojedinim je mjestima došlo do odumiranja posidonije, možda zbog sidrenja ili lova dinamitom pa se razvio Facijes mrtvih naslaga rizoma posidonije bez epiflore (G.3.5.1.3.). Na nekim je područjima kao npr. s južne strane Budikovca, ovaj facijes naseljen invazivnom algom Caulerpa racemosa var. cvlindracea.
Naselja posidonije smatraju se područjima najveće biološke raznolikosti Sredozemnog mora, a iznimno su ugrožena zbog sporog rasta i obnavljanja. Na istraživanom području je utjecaj na posidoniju znatan na mjestima sidrenja kao što su uvale Porat, Mezuporat i Balun te uz Budikovac (Slika 34). Zakonom zaštićeni organizmi zabilježeni u ovim biocenozama:
1. morska cvjetnica posidonija Posidonia
oceanica
2. školjkaš plemenita periska Pinna nobilis
3. puž bačvaš Tonna galea


BOX 2: Naselje posidonije u uvali Rukavac
Gornji rub naselja posidonije u uvali Rukavac nalazi se na 5 metara, dok se donji rub, koji je isprekidan i regresivan, nalazi na 34 metra dubine . Naselje je u većem dijelu kontinuirano, osim na pojedinim mjestima, a posebice u sjevernom i zapadnom dijelu uvale, gdje je naselje krpasto te su zabilježena oštećenja uzrokovana sidrenjem. Porastom dubine i smanjenjem intenziteta svjetlosti, gustoća izdanaka naselja u uvali Rukavac se smanjuje s 384 izdanka po m2na 5 m dubine na 54 izdanka po m2 na 32 m dubine.
Tablica 1. Gustoća naselja posidonije u uvali Rukavac na različitim dubinama (n=3).
| dubina (m) | gustoća (čuperak m’2) |
| 5 | 384 |
| 15 | 252 |
| 20 | 163 |
| 34 | 54 |
Prosječne duljine adultnih listova kreću se od 21,39 cm na 15 m dubine do 34,81 cm na 32 m. Prosječne širine adultnih listova kreću se od 0,74 cm na 5 m do 0,94 cm na 32 m) dubine. Vrijednosti indeksa lisne površine kreću se od 21120 cm2/m2 na 15 m do 5743 cm2/m2na 32 m dubine. Vrijednosti indeksa lisne površine uglavnom su niže od minimalnih vrijednosti za Sredozemno more (20000 cm2/m2). Lepidokronološkom analizom utvrđenje prosječan broj listova po izdanku za posljednjih 15 godina koji se kreće od 2,3 do 7 te je približno jednak za sve četiri dubine.
Vrij ednost koeficij enta A (postotak listova oštećenog vrha) je najmanja na 5 m dubine (0,25%), dok je najveća na 32 m dubine (14,63%) što je u skladu s vrijednostima u Sredozemnom moru.
Iz zračnih snimaka jasno se vidi gornji rub naselji posidonije u uvali Rukavac.
Najveća biomasa po izdanku zabilježena je na 15 m dubine (0,4 g suhe mase) i smanjuje se porastom dubine (0,2 g suhe mase na 32 m). Zbog velikih razlika u gustoći izdanaka po m2, biomasa po površini se također smanjuje (132,8 g suhe mase/ m2 na 5 m do 10,4 g suhe mase/m2 na 32 m). Primarna produkcija po izdanku se smanjuje porastom dubine s 2,7 g suhe mase na 15 m do 0,4 g suhe mase na 32 m dubine te je uglavnom veća od prosjeka zabilježenog za Sredozemno more. Primarna produkcija po m2 također se smanjuje od 957,7 g suhe mase po m2 na 5 m do 24,2 g suhe mase po m2 na 32 m dubine.

G.3.6. Infralitoralna čvrsta dna i stijene
G.3.6.1. Biocenoza infralitoralnih alga
G.3.6.1.1. Degradirani facijes s inkrustrirajućim algama
G.3.6.1.2. Asocijacija s vrstom Cystoseira amantacea var. spicata
Ova biocenoza zahvaća značajnu površinu na istraživanom području (Slika 36). Donju granicu širenja zone prekida naselje posidonije ili strma i zasjenjena obala pa dno i zajednice imaju scijafilni karakter. U najvišoj zoni ove biocenoze često se razvija alga Cystoseira amantacea var. spicata odnosno asocijacija s vrstom Cystoseira amantacea var. spicata (G.3.6.1.2.) (Slika 37). Posebno je dobro razvijena s istočne strane otočića u SE zoni istraživanja te uz istočnu i zapadnu stranu otoka Biševa.
Stjenovitim infralitoralom dominiraju alge, osim u slučajevima kada brojni ježinci dovedu do značajnog golobrsta. Tada se razvija degradirani facijes s inkrustrirajućim algama (G.3.6.1.1.) koji je na istraživanom području zabilježen sporadično između 1 i 5 m dubine. Eksplozije ježinaca objašnjavaju se prelovom njihovih predatora i to riba poput šarga Diplodus sargus i morskih zvjezdača koje se sakupljaju zbog suvenira. U sklopu projekta Pag – Konavle prati se stanje ježinaca na jugozapadnoj strani otoka Ravnika (Slika 38).
Neke svojte alga sunanekimporučj ima, ali i sezonski boljerazvij enepadominiraj uodređenimpodručj ima. Posebno tijekom ljeta, prevladava smeđa alga Padinapavonica na plitkim osvijetljenim mjestima (Slika 39), Acetabularia acetabulum u plitkim zaštićeni]’im mjestima, Coralina spp. u plitkim zasjenjenim te Flabelia petiolata i Halimeda tuna na dubljim zasjenjenim mjestima. Dublja područja ove zajednice naseljavaju invazivne vrste Caulerpa racemosa var. cylindracea i Womersleyella setacea (Slika 40 i 41). Od životinja se posebno ističu zvjezdače Ophidiaster ophidianus, Echinaster sepositus, Hacelia atenuata i Peltaster placenta.
Zakonom zaštićeni organizmi zabilježeni u ovim biocenozama:
1. alga Cystoseira amantacea var. spicata
2. alga Cystoseira spinosa
3. venerin puž Eros aria spurca
4. školjkaš prstac Lithophaga lithophaga
5. trpovi Holothuroidea

Slika 37. Asocijacija s vrstom Cystoseira amantacea var. spicata razvija se u najvišem području infralitoral. Ovaj pojas za vrijeme jakih oseka može u potpunosti biti izvan mora.
6. hridinski ježinac Paracentrotus lividus
7. igličasti ježinac Centrostephanus longispinus
8. zmijolika zvijezda Ophidiaster ophidianus



G.3.8.6. Infralitoralne zajednice s invazivnim vrstama
G.3.8.6.2. Zajednice s vrstom Caulerpa racemosa
Na istraživanom području su zabilježene dvije svojte invazivnih alga: Caulerpa racemosa var. cvlindracea i Womersleyella setacea (Slika 40, 41). Obje su alge na istraživanom području gotovo posvuda rasprostranj. Caulerpa racemosa var. cvlindracea se razvija od same površine do dubina od preko 40 m u biocenozi infralitoralnih alga, naselju posidonije, biocenozi sitnih ujednačenih pijesaka, koraligenskoj biocenozi i biocenozi obalnih detritusnih dna. Womersleyella setacea razvija se od 15 do 30 (40) m dubine u biocenozi infralitoralnih alga, naselju posidonije i koraligenskoj biocenozi. Caulerpa racemosa se pored alge Womersleyella setacea smatra jednom od 10 najinvazivnijih alga u Sredozemnom moru. Nažalost, ne postoje načini njihove kontrole i smanjivanja brzine širenja, a njihov negativni utjecaj s vremenom će se povećavati.

G.4. Cirkalitoral
Cirkalitoralno područje obilježava scijafilna morska vegetacija. Na istraživanom području ono se proteže od oko (20) 30 metara pa do 90 m dubine, što su najveće izmjerene dubine zahvaćene istraživanjem.
Na zasj enj enim mj estima se cirkalitoralne biocenoze razvijaju na plićem dnu pa na dubini od dvadesetak metara nalazimo koraligensku biocenozu kao kod rta Kobila na sjevernoj strani Biševa.
Koraligenska biocenoza posebno je značajna zajednica cirkalitoralnog područja. Biocenoza je dobila ime po crvenim algama porodice Corallinaceae koje ugrađujući kalcijev karbonat u svoje taluse, razvijaju biogene nakupine te čine osnovu ove zajednice. Ovo je područje visoke biološke raznolikosti, a u kojem žive i neki od ekonomski najvrjednijih organizama poput crvenog koralja. Zbog iznimne vizualne šarolikosti, koraligenska biocenoza je posebno zanimljiva roniocima, a posebno na mjestima gdje dominiraju rožnati koralji (Paramuricea clavata, Eunicella spp.). Koraligenska biocenoza je u Hrvatskoj uvrštena u ugroženi tip staništa.
G.4.2. Cirkalitoralni pijesci
G.4.2.2. Biocenoza obalnih detritusnih dna
Horizontalna dna na dubinama između 35 i 90 m dubine naseljava biocenoza obalnih detritusnih dna (G.4.2.2.).

Posebno je lijepo razvijena na zapadnoj strani otoka Biševo gdje započinje na dubini od oko 30 m i blago se spušta prema dubini (Slika 42). Ovo je dno prepuno nepričvršćenih kalcificiranih i mekanih scijafilnih svojti crvenih alga, mahovnjaka, mješčićnica, i mnogočetinaša. Na južnoj strani Biševa unutar ove biocenoze je razvijeno naselje vrste Eunicella singularis (Slika 43).
Biocenoza je ugrožena pridnenim mrežama koje mogu dovesti do oštećivanja mahovnjaka i gorgonija. Sirenje invazivne alge Caulerpa racemosa var. cylindracea ozbiljna je prijetnja plićim područjima ove zajednice.

G.4.3. Cirkalitoralna čvrsta dna i stijene
G.4.3.1. Koraligenska biocenoza
G.4.3.1.5. Facijes s vrstom Eunicella cavolinii G.4.3.1.6. Facijes s vrstom Eunicella singularis G.4.3.1.8. Facijes s vrstom Paramuricea clavata G.4.3.1.9. Facijes s vrstom Parazoanthus axinellae
Zbog konfiguracije morskog dna, čestih podmorskih stonaca, koraligenska biocenoza je značajno rasprostranjena posebno na području Biševa gdje se javlja u obliku facijesa s gorgonijama (Slika 44, 45). Međutim, u dvodimenzionalnoj projekciji karte staništa, površina ove biocenoze je razmjerno mala premda vertikalni strmci u naravi predstavljaju značajnu površinu morskoga dna.
Dobro razvijene facijese s gorgonijama nismo pronašli u SE zoni. Razlog tomu su vjerojatno slabije morske struje.
U koraligenskim biocenozama posebno su dobro razvijeni mahovnjaci čije kolonije ponegdje zauzimaju kompaktne površine od više od jednog kvadratnog metara.
Koraligenska je biocenoza iznimno atraktivno područje za ronilački turizam. Također, ovdje se zadržavaju komercijalno zanimljive vrste pa se na koraligenskim dnima često lovi (Slika 46 i 47). Upravo je lov pridnenim mrežama glavni čimbenik
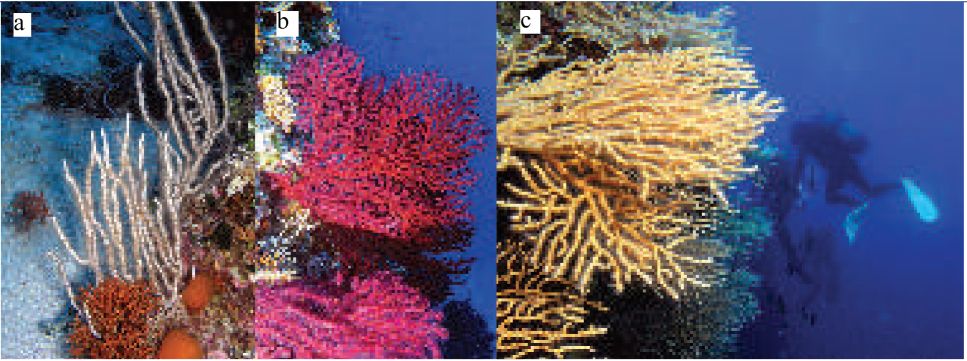
uništavanja koraligena. Ronilački turizam također može dovesti do uništavanja naselja gorgonija slučajnim oštećivanjem zadruga. Invazivna alga Caulerpa racemosa var. cvlindracea također negativno djeluje na ovu biocenozu (Slika 48). Zbog svega toga možemo reći da je koraligenska biocenoza na istraživanom području ugrožena.
Zakonom zaštićeni organizmi zabilježeni u ovoj biocenozi:
1. igličasti ježinac Centrostephanns longispinus
2. rak jastog Palinurus elephas
3. žuta gerardia G er ardi a s avaglia.


l\/Tr\Hrp*



Inventarizacija vrsta
Inventarizacija vrsta istraživanog područja obuhvaća fitobentos, zoobentos, fitoplankton, zooplankton, riblje vrste i sisavce. Riblje vrste date su u izvještaju Zadataka 2.
Ukupno je zabilježeno 181 svojta makroalga, 2 vrste cvjetnice, 269 beskralješnjaka, 347 fitoplanktonskih i 100 zooplanktonskih svojti te 4 vrste kitova. Uz 114 vrsta riba zabiljženih na širem području Viškog arhipelaga, ukupno je zabilježeno 1017 svojti (Prilozi 1 – 5).
Na istraživanom je području tijekom istraživanja zabilježeno 19 zaštićenih svojti:
14. trp Holothuria forskali
15. trp Holothuria tubulosa
16. spužva špiljska sumporača Aplysìna cavernicola
17. rak jastog Palinurus elephas
18. žuta gerardia G er ardi a s avaglia
19. dobri dupin Tursìops truncatus
Prijašnjim istraživanjima utvrđene je za šire područje Visa 10 zaštićenih svojte, a koje sada nisu pronađene:
1. morska naranča Tethva aurantium
2. periska Atrina fragilis
3. tritonova truba Charonia tritonis
4. hlap Homarus gammarus
5. rakovica Maja squinado
6. kuka Scyllarides latus
7. zezavac Scyllarus sp.
8. trp Eostichopus regalis
9. trp Holothuria sanctori
10. trp Ocnus planci
1. morska cvjetnica posidonija Posidonia oceanica
2. morska cvjetnica čvorasta morska resa Cymodocea nodosa
3. alga Titanoderma trochanter
4. alga Cystoseira amantacea var. spicata
5. alga Cystoseira spinosa
6. puž zupka Luria lurida
7. venerin puž Prosaria spurca
8. bačvaš Tonna galea
9. školjkaš prstac Lithophaga lithophaga
10. školjkaš plemenita periska Pinna nobilis
11. hridinski j ežinac Paracentrotus lividus
12. igličasti ježinac Centrostephanus longispinus
13. zmijolika zvijezda Ophidiaster ophidianus
Korištenje obale i mora
Područje obale i mora istraživanog područja iskorištava se u rekreativne, gospodarske i prometne svrhe (Slika 49).
Obala, posebno pješčana i šljunčana, koristi se za plaže, naročito tijekom ljetnog razdoblja. Koristi se još i za izvlačenje brodica i njihov suhi vez. Stjenovita obala je na istraživanom području relativno strma, pa se kao takva ne iskorištava kao plaža. Obala i more koriste se za rekreaciju, sunčanje, kupanje i ronjenje na dah.
U uvalama je dio obale betoniziran kako bi se napravila pristaništa za brodice.
Morske špilje se koriste u turističke svrhe. Postoje i organizirani obilasci Zelene, Medvidove i Modre špilje. Modra špilja je pod koncesijom pa je za ulazak u nju potrebno platiti ulaznicu.
More se koristi kao plovni put rekreativnih i transportnih brodica. Jedini trajno organizirani transport povezuje Komižu s otokom Biševo odnosno uvalama Mezuporat, Salbunara i Porat. Većina uvala koristi se za sidrišta, a u naseljenim mjestima postavljeni su i pridneni blokovi za privez brodica. U uvali Rukavac postavljeno je nekoliko plutača za privez nautičkih brodova, a u uvali Mezuporat privezište je organizirano preko sidrenih blokova povezanih murinzima za betonsku obalu.
Podmorje se iskorištava za ribolov i to sportski, dopunski, gospodarski, znanstveni i nezakoniti. Podmorje se iskorištava i za autonomno ronjenje i to rekreativno, znanstveno i profesionalno. Rekreativno autonomno ronjenje moguće je jedino preko ronilačkih centar koji za ovo područje imaju koncesiju. Ukupno su četiri ronilačka centra i to dva u Visu i dva u Komiži.
Na SE zoni istraživanja gotovo da nema ronilačkih lokaliteta u kategoriji sportskog ronjenja na koje lokalni ronilački centri vode ronioce. Jedina područja, ali s malim intenzitetom ronjenja su Zelena špilja i otočić Greben. Na području Biševa je nekoliko lokaliteta za organizirano ronjenje i to rt Gatula, Modra špilja, hrid Totac i rt Kobila. Ipak, centri iz Komiže češće vode ronilačke ture na lokalitete oko rta Stupišće i rta Barjak. Ovi su lokaliteti iznimno zanimljivi zbog koraligenske biocenoze i naselja gorgonija te arheoloških nalaza. Centri iz Visa vode ronioce na lokalitete neposredno ispred viške luke koji su zanimljivi također zbog koraligenske biocenoze i naselja gorgonija te arheoloških nalaza.
Nismo pronašli vođene ture koje nude ronjenja s maskom i disalicom, a posebno uz interpretacijska vođenja, što bi posebno moglo biti zanimljivo za područje juže strane Biševa, gdje postoji izniman potencijal iskorištavanja prostora u turističke svrhe. Osim podmorja i posebnog krajobraza, moguće je vidjeti i rijetke crne sokolove koji se ovdje gnijezde. Također je moguće organizirati posjet Medvidovoj pećini uz korištenje maske, disalice i podvodne lampe što bi turistima pružilo jedinstven uvid u ekosustave morskih pećina i nezaboravan doživljaj.

Ugroze biološke i krajobrazne raznolikosti obale i mora
Istraživano područje u osnovi je dobro sačuvano i bez značajnijih ugroza. Ipak, pojedina staništa su pod većim pritiskom ljudskog djelovanja ili invazivnih vrsta (Slika 50).
Pj eskovita i šlj unčana obala se tij ekom ljeta koristi za plaže pa su u to vrijeme ova staništa pod značajnim ljudskim pritiskom posebno zbog gaženja. Tijekom jeseni, zime i proljeća, ovdje se nakuplja značajna količina antropogenog naplavljenog otpada premda on u principu nema značajniji negativni efekt. Izvlačenje brodova i njihov suhi vez također minimalno utječe, ali skidanje antiobraštajnih boja s brodova i njihovo nakupljanje u detritusu može negativno djelovati na organizme.
Obalna gradnja je do sada bila minimalna, ali i daljnja gradnja, također, mora biti minimalna. Neadekvatno postavljanje sidrenih lančanika kod trajnih privezišta na nekim mjestima djeluje negativno na morsku cvjetnicu Posidonia oceanica kao što je slučaj u uvalama Mezuporat i Rukavac. Na naselja posidonije negativno djeluje i slobodno sidrenje koje je posebno intenzivno oko otoka Budikovca, te u uvalama Rukavac, Mezuporat i Balun.
Ribolov pridnenim mrežama osim na osiromašenje ciljanih komercijalnih vrsta, negativno djeluje i na neke pridnene zajednice, posebno koraligen, jer dovodi do fizičkog uništavanja koraligenskih vrsta. Ronilački turizam djeluje negativno ukoliko ronioci oštećuju ili uznemiruju organizme. Potrebno je napomenuti daje ronjenje moguće isključivo preko ovlašteni ronilačkih centara. Međutim, za vrijeme ronjenja nije potrebno da u pratnji ronioca bude i netko iz ronilačkog centra. Ovo je apsurdno jer na ovaj način ne postoji nikakva kontrola onoga što ronioci u podmorju rade!
Intenzivan plovni promet, posebno bučni brodovi kao što su crusing brodovi, a koji plove između Visa i Biševa, stvaraju podmorsku buku koja utječe na dupine.
Na istraživanom su području zabilježene dvije invazivne alge: Caulerparacemosavar. cylindracea i Womersleyella setacea. Obje su alge široko rasprostranjene, a njihovo uklanjanje nije moguće. Canlerpa racemosa var. cylindracea se razvija od same površine do dubina od preko 40 m u biocenozi infralitoralnih alga, naselju posidonije, biocenozi sitnih uj ednačenihpij esaka, koraligenskoj biocenozi i biocenozi obalnih detritusnih dna.
Womersleyella setacea razvija se od 15 do 30 (40) m dubine u biocenozi infralitoralnih alga, naselju posidonije i koraligenskoj biocenozi.

Posebno odabrana područja
Uzimajućiuobzirpodručja veće biološke raznolikost staništa i vrsta, potencijalom iskorištavanja u turističke svrhe i minimalnog sukoba interesa (prvenstveno s ribarima) odabrana su dva područja za koje je na prezentaciji projekta u Komiži (07. 11. 2008.) predložen poseban režim upravljanja (vidi izvještaj sa prezentacija: Prilog 6). Na ovim bi se područjima uvela „no take“ zona, a područja bi bila korištena u turističko rekreativne svrhe. Ideja „no take“ zone j e prvenstveno u cilju obnove ekosustava, komercijalnih resursa i konačno boljeg ribolova u okolnom području (Vidi također izvještaj Zadatak 2). Ovakva bi zona postala turistički zanimljiva i time bi se otvorila nova dodatna turistička ponuda, ali i dobio brend zaštićenog podmorja.
Oba odabrana područja su na Biševu i to na južnoj i jugoistočnoj strani otoka. Područja su specifična po iznimnoj krajobraznoj zanimljivosti i očuvanosti kopna i podmorja te prisutnosti brojnih biocenoza (Slika 51). Ipak, u diskusiji s ribarima tijekom radionice, premda je općenita ideja o „no take“ zoni podržana, ribari se nisu u potpunosti složili s predloženim zonama jer neke pokrivaju njihova najbolja ribolovna područja. Ribari se slažu da da treba osnovati „no take“ podučje i za njega predlažu jugoistočnu stranu Biševa (vidi izvještaj sa prezentacija: Prilog 6).

Slika 51. Prijedlog info ploče o podmorju Biševa prezentirane na prezentaciji projekta u Komiži (07.11. 2008.), a u cilju promidžbe turizma Viškog arhipelaga.
Preporuke za uobičajeni pristup istraživanju bioraznolikosti morskog područja
Istraživanje bioraznolikosti nekog priobalnog morskog područja uključuje sljedeće faze: ekipiranje
prikuplj anj e postoj ećih podataka terenski rad
laboratorijska analiza uzoraka izrada izvještaja popularna interpretacija
Ekipiranje
Za istraživanje biološke raznolikosti većeg dijela našeg priobalnog podmorja, u većini slučajeva će se pažnja posvetiti bentoskim organizmima i ribljim zajednicama. Bentoska istraživanja dijelimo na zoobentoska i fitobentosa. Kod zoobentoskih organizama uglavnom se pažnja posvećuje mekušcima, žamjacima, spužvama, mnogočetinašima i mahovnjacima. Za što bolji rezultat, u ekipi bi za svaku od ovih skupina trebao postojati specijalizirani taksonom. Kod fitobentosa se se pretpostavlja da fitobentoničar pokriva smeđe, crve i zelene alge te morske cvjetnice. Istraživanje modrozelenih alga zahtjeva specijaliziranog taksonoma.
Kod ihtioloških istraživanja se pretpostavlja da su istraživanja adultnih i juvenilnih stadija dvije odijeljene specijalnosti koje pokrivaju različiti stručnjaci.
Kod specifičnih staništa kao što su morska jezera, javljaju se i druge skupine organizama koje treba pokriti istraživanjem kao npr. zooplankton i fitoplankton. U ovakva staništa potrebno je uključiti i fizikalno kemijske parametre jer su oni osnova za razumijevanje takvih ekosustava.
U cilju izrade G1S karata potrebno je imati posebni tim sposoban za terensko prikupljanje podataka (npr. batimetrija), te uredsku obradu podataka.
Prikupljanje postojećih podataka
Postojeći se podaci prikupljaju iz objavljenih stručnih i znanstvenih radova koji su obrađivali istraživano ili blisko područje. Činjenica je da je mnoštvo sakupljenih podataka još uvijek neobjavljen, a do takvih je podataka teško doći ili zahtijevaju značajan napor kako bi se obradili.
Terenski rad mora biti sezonski. On uključuje sakupljanje uzoraka za laboratorijsku analizu, in situ popisivanje organizama, fotografiranje i snimanje. Metodologije sakupljanje uzoraka ovisi o ciljanoj istraživanoj skupini. Često uključuje autonomno ronjenje, korištenje ribolovnih alata i specijaliziranih alata za sakupljanje uzoraka kao stoje grabilo ili dredža.
Terensko istraživanje treba se planirati barem kroz dvije godine kako bi se pokrile sezonske vrste i godišnja odstupanja u njihovom pojavljivanju.
Laboratorijska analiza uzoraka
Laboratorijska analiza uzoraka dugotrajan je i specijalistički proces. Nije čudno ako za neka taksonomska istraživanja određenih skupina treba više mjeseci ili godina kako bi se analizirali uzorci jednog terena. Npr. smatra se da za analizu jednog uzorka npr. s površine od 20 za 20 cm) makrobentosa (uključuje fitobentos i zoobentos) potrebno je minimalno jedan radni dan uz sudjelovanje svih specijalista za pojedine bentoske skupine. Da bi se analizirao bentos jednog transekta potrebno je sakupiti i analizirati dubinske uzorke. Oni su obično raspoređeni na dubinske razrede i to: supralitoral, mediolitoral, 0-0,5m, 0,5-lm, l-5m, 5-10m, 10-20, 20-30, 30-40, dublje od 40m (obično dva uzorka dredžom ili grabilom). Prema tome, smatra se da za analizu samo jednog transekta bentosa treba barem 10 dana intenzivnog laboratorijskog rada uz sudjelovanje minimalno 2 specijalista (fitobentoničar i zoobentoničar).
Upravo zbog sporosti laboratorijske analize bentoskih i planktonskih organizama, za ukupna taksonomska istraživanja nekog odabranog područja, a kako bi se kvalitetno obradile sve sezone, potrebno je najmanje dvije godine.
Izrada izvještaja
Izvještaj mora prikazati postojeće stanje na terenu kroz pregled, zatečenih staništa (G1S pregled), vrsta (inventome liste), analizu ugroza, korištenja prostora, preporuke za očuvanje biološke raznolikosti ili obnovu oštećenih ekosustava.
Popularna interpretacija bitna je iz više razloga:
– kako bi se lokalnom stanovništvu prikazale vrijednosti njihovog podmorja, ukazalo na mogućnosti očuvanja i održivog korištenja;
– kako bi se promoviralo područje u turističke svrhe
– kako bi se utjecalo na opću svijest u cilju zaštite.
Najčešći načini popularne interpretacije su web stranice, info ploče, posteri i brošure i promotivni filmovi. Izrada ovih materijala uključuje niz profesionalaca. Podvodni fotografi i snimatelji, web dizajneri, grafičari, montažeri i si. Uz njih je potreban i biolog koji će dati osnovni koncept, pripremiti interpretaci]’ske tekstove, objašnjenja, znanstvene nazive i si.
Troškovnik za uobičajeni pristup istraživanju bioraznolikosti morskog područja
Cijena realizacije cijelog procesa istraživanja biološke raznolikosti morskog područja do popularne interpretacije, ovisi o mnoštvu specifičnih čimbenika i gotovo je nije moguće teoretski predvidjeti.
Tako npr. ovisi o mjestu istraživanja: udaljeni otok ili priobalno područje. Udaljeni otok iziskuje posebnu logistiku (prijevoz, smještaj). Na terensko istraživan] e izdvoj enih otoka više utj eče loše vrij eme pa je obično potrebno utrošiti i više terenskih dana. Pitanje je dali se koristi automobil, manja brodica ili istraživački brod, a sve može drastično promijeniti cijenu.
Okvirni proračun za 7 dana terenskog rada, 5 istraživača, kapetan brodice, korištenje manje brze brodice, automobila, smještaj, prijevoz, materijalni troškovi i naknade za ronjenje trebao bi iznositi minimalno 75000,00 kn. Tijekom sedam dana terena uz idealno vrijeme, realno je očekivati da će se obraditi 5 profila (postaja).
Koliko transekata treba obraditi po istraživanom području nije moguće teoretski odrediti. Npr. za preliminarno istraživanje bentosa Biševa bilo bi potrebno minimalno 5 transekata tijekom barem dvije sezone. Za laboratorijsku analizu sakupljenih uzoraka s pet transekata (ukupno oko 50 uzoraka) bilo bi potrebno oko 50 dana uz minimalno sudjelovanje 2 specijalista.
Realna cijena ovog posla je 150.000,00 kn. Kako u analizu nekog područja treba uključiti i ihtiologe te stručnjake specijaliste za pojedine taksonomske skupine, cijena prelazi 200.000, 00 kn za analizu jednog terena.
U cijenu finalne izrade izvještaja treba uzeti u obzir i izradu G1S karata na kojima će biti prezentirani zabilježena staništa, rijetke vrste, invazivne vrste, batimetrija i si.
Za izradu batimetrije potrebna su dodatna terenska mjerenja. Za izradu G1S karata potrebno je kupiti ortofoto snimke.
Izrada finalnihproizvodazapopulamu interpretaciju uključuje sudjelovanje mnogih profesionalaca. Prosječna cijena podvodne fotografije je oko 400 kn. Cijena fotografija je daleko veća ako je za fotografije potrebno otići na teren kako bi se dobile specifične fotografije, npr. iz Medvidove spilje na Biševu.
Grafička izvedba prosječne brošure kreče se od minimalno 4000 kn. Potrebnoj e platiti osmišlj avanj e brošure i pisanje popratnog tekstualnog dijela (2000,00 kn) te izradu crtanih dijelova brošure kao što su sheme, dubinski profili i si (2000,00 kn). Ako brošura sadrži minimalno 20 fotografija, njena cijena bi bila oko 16000,00 kn.
Prema tome, cijena preliminarnog istraživanja bentosa i ribljih zajednica priobalnog područja kao što je područje Biševa (terenski rad tijekom dvije sezone, laboratorijska obrada, izvještaj i priprema tri brošure) bila bi minimalno 650.000,00 kn.
Preporuke za daljnje aktivnosti u Viškom akvatoriju
Daljnjim aktivnostima potrebno je nadopunjavati spoznaje istraživanih područja (Biševo, SE Vis), ali i proširiti istraživanja na šire područje Viškog arhipelaga. Ronilačke brošure koje su pripremljene za tisak kao i info ploču, potrebno je tiskati i razdijeliti odnosno postavi info ploču u dogovoru sa zainteresiranim strankama (Grad Vis i Komiža, turističke zajednice, ronilački centri) (Slika 52). Potrebno je podupirati nastojanja da se unaprijedi promidžba Visa kao turističke destinaci]’ e s brendom područja visoke biološke raznolikosti. S tim ciljem može se poduprijeti izrada dokumentarnog filma o biološkoj raznolikosti podmorja Viškog akvatorija.
U suglasnosti sa zainteresiranim strankama treba raditi na provedbi ideje o osnivanju posebno upravljanog područja s „no take“ zonom te svim što uz to ide.
Dodatne aktivnosti na područja Biševa i SE Visa trebaju obuhvatiti:
detaljnije kartiranje naselja gorgonija praćenje stanja trotoara i alge Cystoseira amantacea var. cpicata osnivanje posebno upravljanog područja (no take zone) te slijedom toga:
a) detaljnu analizu bentoskih i ihtio naselja
upravljanog područja prije početka provedbe
b) praćenje i izvještavanje o provedbi
upravljanog područja (na kraju svake naredne godine)
c) praćenje i izvještavanje o promjenama bentoskih i ihtio naselja upravljanog područja (na kraju svake naredne godine)
d) javna prezentacija provedbe i senzibilizacija
javnosti (stalno)
Daljnje aktivnosti na širem području Viškog akvatorija trebaju obuhvatiti:
– oređivanje hot spot mjesta biološke raznolikosti ili rijetkih, zaštićenih, specifičnih svojti ili staništa
– obradu odabranih hot spot područja sukladno Preporukama za uobičajeni pristup istraživanju bioraznolikosti morskog područja.
– istraživanja dupina
– izradu promidžbenog materijala (dokumentarni filmovi, web stranice, info ploče, brošure i si).
PRILOG 1. Fitobentos
Inventoma lista alga i morskih cvjetnica područja Biševa i SE Visa načinjena je na osnovu dosadašnjih neobjavljenih podataka sastava flore otoka Biševa i područja Rukavca, te podataka dobivenih ovim istraživanjem. Ukupno je zabilježeno 183 svojte (Tablica 1 i 2).
Tablica 1. Ukupna brojčana (N) i postotna (%) zastupljenost odjeljaka bentoskih alga i morskih cvjetnica na otoku Biševu i područja SE Visa.
| Taksonomski odjeljak | N | % |
| Rhodophyta | 121 | 66,1 |
| Ochrophyta (Phaeophyta) | 35 | 19,1 |
| Chlorophyta | 25 | 13,7 |
| Tracheophyta (Spermatophyta) | 2 | U |
| UKUPNO | 183 |
Tablica 1. Bentoske alge i morske cvjetnice na području otoka Biševa i SE Visa.
I RACHEOPHYTA (SPERMATOPHYTA) – MORSKE CVJETNICE
Cymodocea nodosa (Ucria) Acherson Posidonia oceanica (Linnaeus) Delile
CHLOROPHYTA – ZELENE ALGE
Acetabularia, acetabulum (Linnaeus) P.C. Silva ‘ Anadyomene stellata (Wulfen) C.Agardh
Bryopsis hypnoides J.V.Lamouroux Caulerpa racemosa (Forsskâl) J.Agardh I Chaetomorpha aerea (Dillwyn) Kiitzing
i Chaetomorpha mediterranea (Kiitzing) Kiitzing var. crispa (J.Feldmann) (Feldmann) Gallardo et al.
Cladophora coelothrìx Kiitzing Cladophora dalmatica Kiitzing I Cladophora pellucida (Hudson) Kiitzing
Cladophora prolifera (Roth) Kiitzing Codium bursa (Linnaeus) C.Agardh Codium effusum (Rafinesque) Delle Chiaje Dasycladus vermicularis (Scopoli) Krassar Enteromorpha multiramosa Bliding Entocladia endolithica (Ercegovic) R.Nielsen I Flabellia petiolata (Turra) Nizamuddin
i Halimeda tuna (J. Ellis et Solander) J.V. Lamouroux Palmophyllum crassum (Naccari) Rabenhorst Pedobesia simplex (Meneghini ex Kiitzing) M.J. Wynne et Leliaert I Phaeophila dendroides (P.Crouan et H.Crouan) Batters
Pseudochlorodesmis furcellata (Zanardini) Borgesen Pseudochlorodesmis tennis Ercegovic . Rhizoclonìum tortuosum (Dillwyn) Kiitzing
Valonia macrophysa Kiitzing
Vnlnmin ntrimìnri? tR nitit ( AaçtrrWI
Acrochaetium virgatulum (Harvey) Batters Acrosymphytonpurpuriferum (J.Agardh) G. Sjöstedt Acrosorium ciliolatum (Harvey) Kylin
Aglaothamnion tenuissimum (Bonnemaison) Feldmann-Mazoyer Anotrichium barbatum (C. Agardh) Nägeli
Antithamnion cruciatum (C.Agardh) Nägeli var. profundum Feldmann Mazoyer
Antithamnion heterocladum Funk Antithamnion tenuissimum (Hauck) Schiffner Antithamnionella spirographidis (Schiffner) E.M. Wollaston Amphiroa beauvoisii J.V.Lamouroux Amphiroa cryptarthrodia Zanardini Amphiroa rigida J.V.Lamouroux Apogiossum ruscifolium (Turner) J.Agardh Balliella cladoderma (Zanardini) Athanasiadis Boergeseniella fruticulosa (Wulfen) Kylin Botryocladia botryoides (Wulfen in Jacquin) J.Feldmann Botryocladia chiajeana (Meneghini) Kylin Botryocladia microphysa (Hauck) Kylin Callithamnion corymbosum (J.E. Smith) Lyngbye Catenella caespitosa (Withering) L.M. Irvine Caulacanthus ustulatus (Martens ex Turner) Kützing Ceramium bertholdii Funk Ceramium ciliatum (J.Ellis) Ducluzeau Ceramium codii (H.Richards) G.Mazoyer Ceramium diaphanum (Lightfoot) Roth Ceramium deslongchampii Chauvin ex Duby Ceramium flaccidum (Harvey ex Kützing) Ardissone Ceramium siliquosum (Kützing) Maggs et Hommersand Champiapannila (C.Agardh) Harvey Chondria capillaris (Hudson) M.J. Wynne Chondria dasyphylla (Woodward) C. Agardh Chondrophycus papillosus (C. Agardh) Garbary et Harper Chylocladia verticillata (Lightfoot) Bliding Colaconema daviesii (Dillwyn) Stegenga Compsothamnion thuyoides (J.E. Smith) Nägeli Corallina officinalis Linnaeus
Corallophila cinnabarina (Grateloup ex Bory de Saint-Vincent) R.E. Norris Crouania attenuata (C.Agardh) J.Agardh Dasya baillouviana (S.G. Gmelin) Montagne Dasya hutchinsiae Harvey
Dasya corymbifera J.Agardh Dasya ocellata (Grateloup) Harvey
Dipterosiphonia rigens (Schousboe ex C.Agardh) Falkenberg Dudresnaya verticillata (Withering) Le Jolis Erythroglossnm sandrianum (Zanardini) Kylin Eupogodonplanus (C.Agardh) Kiitzing Eupogodon spinellns (C.Agardh) Kiitzing
Falkenbergia rnfolanosa (Harvey) F. Schmitz – tetrasporofit od Asparagopsis armata Harvey
Fauchea repens (C.Agardh) Montagne et Bory
Gastroclonium clavatum (Roth) Ardissone
Gelidium crinale (Hare ex Turner) Gaillon
Gelidium spinosum (S.G. Gmelin) RC. Silva
Gelidium spathulatum (Kiitzing) Bomet
Griffithsia phyllamphora J.Agardh
Griffithsia schousboei Montagne
Gulsonia nodulosa (Ercegović) Feldmann et G. Feldmann-Mazoyer
Gymnothamnion elegans (Schousboe ex C.Agardh) J.Agardh Halymenia elongata C. Agardh
Halymeniafloresia (Clemente) C.Agardh var. ulvoidea Codomier
Halymneia mucosa Ercegović
Haliptilon virgatum (Zanardini) Garbary et
Halydictyon mirabile Zanardini
H. secunda (C. Agardh) Ambronn f. secunda
H. secunda (C. Agardh) Ambronn f. tenella (C.Agardh) MJ.Wynne
Heterosiphonia crispella (C. Agardh) M.J. Wynne
Hydrolithon farinosum (J.V. Lamouroux) D. Penrose et Y.M. Chamberlain
Hypnea musciformis (Wulfen) J.V. Lamouroux
Hypoglossum hypoglossoides (Stackhouse) F.S. Collins et Hervey
Ja nia rubens (Linnaeus) J.V.Lamouroux
Laurencia obtusa (Hudson) J.V.Lamouroux
Lejolisia mediterranea Bomet
Liagora viscida (Forsskâl) C.Agardh
Lithophyllum byssoides (Lamarck) Foslie
Lithophyllum incrustans Philippi
Lithophyllum racemus (Lamarck) Foslie
Lithophyllum stictaeforme (J.E. Areschoug) Hauck
Lomentaria . chylocladiella Funk
Lophosiphonia cristata Falkenberg
Meredithia microphylla (J.Agardh) J.Agardh = Kallymenia microphylla J. Agardh Monosporus pedicellatus (Smith) Solier var. pedicellatus
Monosporus pedicellatus (Smith) Solier var. tenuis (Feldmann-Mazoyer) Huisman et Kraft
Myriogramme tristromatica (J.J.Rodriguez ex Mazza) Boudouresque
Nemalion helminthoides (Velley in Withering) Batters
Nitophyllum punctatum (Stackhouse) Greville
Osmundaria volubilis (Linnaeus) R.E. Norris
Osmundea pinnatifida (Hudson) Stackhouse
Parviphycus tenuissimus (Feldmann et Hamel) B. Santelices
Peyssonnelia polymorpha (Zanardini) F. Schmitz
Peyssonnelia rubra (Greville) J.Agardh
Peyssonnelia squamaria (S.G. Gmelin) Decaisne
Phymatolithon calcareum (Pallas) Adey et Mc Kibbin
Phymatolithon lenormandii (Areschoug) Adey
Piatoma cyclocolpa (Montagne) F. Schmitz
Plocamium cartilagineum (Linnaeus) P.S. Dixon
Pneophyllum fragile Kützing
Polysiphonia elongata (Hudson) Sprengel
Polysiphonia scopulorum Harvey
Polysiphonia sertiliarioides (Grateloup) J.Agardh
Parviphycus tenuissimus (Feldmann et Hamel) B. Santelices
Ptilothamnion plumula (J.Ellis) Nägeli
Rhodophyllis divaricata (Stackhouse) Papenfus
Rhodymenia ardissonei J.Feldmann
Rodriguezella strafforelloi F. Schmitz ex J.J.Rodriguez var. strafforelloi Rodriguezella strafforelloi F. Schmitz ex J.J.Rodriguez var. crassìcaulis Ercegovié Rytiphlaea tinctoria (Clemente) C.Agardh
Schottera nicaeensis (J.V. Lamouroux ex Duby) Guiry et Hollenberg
Sahlingia subintegra (Rosenvinge) Kommann
Seirospora interrupta (J.E.Smith) F.Schmitz
Spermothamnion flabellatum Bornet
Spermothamnion repens (Dilwin) Rosenvinge
Sphaerococcus coronopifolius Stackhouse
Spongìtes frutìculosus Kützing
Spyridiafilamentosa (Wulfen) Harvey
Stylonema alsidii (Zanardini) K.M. Drew
Titanoderma cystoseìrae (Hauck) Woelkerling, Chamberlain et Silva Titanodermapustulatum (Lamouroux) Nägeli
Titanoderma trochanter (Bory) Benhissoune, Boudouresque, Perret-Boudouresque et Verlaque
Tenarea tortuosa (Esper) M. Lemoine
Tricleocarpa fragilis (Linnaeus) Huisman et Townsend
Womersleyella setacea (Hollenberg) R.E. Norris
Wrangelia penicillata (C.Agardh) C.Agardh
Acinetospora crinita (Carmichael ex Harvey) Sauvageau Ascocyclus orbicularis (J. Agardh) Kjellman Cladosiphon mediterraneus Kützing Cystoseira adriatica Sauvageau
Cystoseira compressa (Esper) Gerlof et Nizamuddin f. rosetta (Ercegović) Cormaci et ai Cystoseira corniculata Hauck ssp. laxior Ercegović Cystoseira crinita Duby Cystoseira spicata Ercegović
Dictyopterispolypodioides (A.P.de Candolle) J.V. Lamouroux Dictyota dichotoma (Hudson) J.V. Lamouroux Dictyota linearis (C.Agardh) Greville Dictyota fasciola (Roth) J.V. Lamouroux Dictyota spiralis Montagne
Ectocarpus siticulosus (Dillwyn) Lyngbye var. adriaticus (Ercegović) Cormaci et G.Fumari Feldmannia caespitula (J.Agardh) Knoepffler-Péguy
Feldmannia irregularis (Kützing ) G. Hamel var. lebeliides (Ercegović) Špan et Antolić
Giraudia sphacelarioides Derbès et Sober
Halopterisfilicina (Grateloup) Kützing
Leathesia mucosa J.Feldmann
Myriactula rivulariae (Suhr) J.Feldmann
Myriactula stellulata (Harvey) Levring
Nereia filiformis (J.Agardh) Zanardini
Padina pavonica (Linnaeus) J.V. Lamouroux
Ralfsia verrucosa (Areschoug) Areschoug
Sargassum vulgare C.Agardh
Spermatochnus paradoxus (Roth) Kützing
Sphacelaria cirrosa (Roth) C.Agardh
Sphacelaria fusca (Hudson) S.F. Gray
Sphacelaria plumula Zanardini
Sphacelaria tribuloides Meneghini
Sporochnuspedunculatus (Hudson) C.Agardh
Stilophora tenella (Esper) P.C.Silva
Stypocaulon scoparium (Linnaus) Kützing
Zanardinia typus (Nardo) RC.Silva
Taonia atomaria (Woodward) J.Agardh
Prilog 2. BESKRALJEŠNJACI
Popis beskralješnjaka je sastavljen na osnovi zabilježenih vrsta tijekom ovih istraživanja, ranije sakupljenih podataka tijekom istraživanja prilova koćarskih lovina šireg područja otoka Visa (neobjavljeni podaci) te dostupnih literaturnih zapisa: Simunović (1991). Ukupno je zabilježeno 269 svojti bentoskih beskralješnjaka od kojih 23 zakonom zaštićene svojte (Tablica 1).
Literatura:
Simunović, A. (1991) Ekološka i biogeografska istraživanja puževa prednješkržnjaka (Gastropoda, Proso-branchiata) u istočnom dijelu Jadranskog mora i njihov odnos prema bentoskim biocenozama. Disertacija, Sveučilište u Zagrebu, Prirodoslovno-matematički fakultet, Zagreb, 283 str.
Tablica 1. Beskralješnjaci Viškog arhipelaga.
Porifera (16 svojti)
Aplysìna aerophoba Aplysina cavernicola Axinella damicornis Crambe crambe Hemimycale columella Hippospongia communis Ircinia muscarum Ircima sp.
Petrosiaficiformis Raspatila viminalis Rhizaxinella pyrifera Sigmatoxella annexa Spirastrella cunctatrix Spongia officinalis lamellosa Tethya aurantium Thenea muricata
Cnidaria (22 svojte)
Actinia equina Alcyonium palmatum Anemonia sulcata Caryophyllia inornata Caryophyllia smithii Ceratotrochus magnaghii Cerianthus membranaceus Condylactis aurantiaca Eunicella cavolinii Eunicella singularis Funiculina quadrangularis Gerardia savaglia Leptopsammia pruvoti Lytocarpia myriophyllum Monomyces pygmaea Paracyathus pulchellus Paramuricea clavata
Pareiythropodium coralloides Pennatula phosphorea Phyllangia mouchezi Plumulariidae sp.
Pteroeides spinosum
Echiurida (1 svojta)
Bonellia viridis
Mollusca (57 svojti)
Alvania cimex Arca noae Astraea rugosa Atrina fragilis Bittium reticulatum Cerithium vulgatum Calliostoma conulum Capulus hungaricus Ceratostoma erinaceum Charonia tritonis seguenza Clanculus cruciatus Columbella rustica Cymathium corrugatum Diodora gibberula Diodora gr aeca Diodora italica Emargìnula huzardi Eros aria spurca Fasciolaria lignaria Flabellìna affinis Fusinus syracusanus Galeodea echìnophora Gastrochaena dubia Gibbula adansoni Gibbula magus Gibbula sp.
Hexaplex trunculus Hinìa incrassata Janolus cristatus Jujubinus striatus Lemìntìna arenaria Lima lima
Lìthophaga lithophaga Luria lurida Melarhaphe nerìtoìdes Muricopsis cristatus Mytilus galloprovìncìalìs Ocìnebrìna aciculata Osilìnus turbinatus Ostrea cochlear Patella rustica Patella ulyssiponensis Pectenjacobaeus Pinna nobilis Pisanìa striata Pteria hirundo Raphitoma reticulata Rissoa sp.
Scaphander lignarius Scaphopoda sp.
Simnia spelta Spondylus gaederopus Susania testudinaria Thuridilla hopei Tonna galea Venus verrucosa Vermetus triqueter
Annelida (32 svojte) Eunice oerstedìi Eunice sp.
Eunice torquata
Eunice vittata Filograna sp.
Glycera rouxiì Harmothoe areolata Harmothoe sp.
Harmothoe spinifera Hermodìce carunculata Hesionidae sp.
Hyalinoecia tubìcola Janua pagenstecheri Leocrates chinensis Lumbrinerìs coccinea Lysidice ninetta Nereis rava Nereis sp.
Pomatoceros trìqueter Protula sp.
Sabellidae sp.
Serpula vermicularis Serpulidae sp. Spirobranchus polytrema Spirographis Spallanzani Spirorbis infundibulum Subadyte pellucida Syllidae sp.
Syllis sp.
Vermilìopsìs infundibulum Vermilìopsis labiata Vermiliopsis striaticeps
Arthropoda (33 svojte) Alpheus dentipes Balanus sp.
Chthamalus depressus Chthamalus stellatus Chthamalus montagui Dromi a personata Eriphìa verrucosa Galathea sp.
Goneplax rhomboïdes Inachus sp.
Homarus gammarus Latreìllia elegans Ligia italica Liocarcinus depurator Lissa chiragra Macropodia longirostrìs Macropipus tuberculatus Maja squinado Maia sp.
Pachygrapsus marmoratus Palemon elegans Palinurus elephas
Parthenope sp.
Pilumnus hirtellus Pisa nodipes Processa nouveli nouveli Scalpellum scalpellum Scyllarides latus Scyllarus sp.
Solenocera membranacea Squilla mantis Stenopus spinosus Xantho sp.
Tentaculata (72 svojte) Adeonella pallasii Aetea sica Aetea truncata Annectocyma sp.
Beania magellanica Caberea boryi Callopora dumerilii Callopora lineata Cellaria salicornioides Celleporina caminata Celleporina lucida Chorizopora brongniartii Copidozoum planum Crassimarginatella crassimarginata Crassimarginatella maderensis Crassimarginatella solidula Cris i a sp.
Diplosolen obelia Disporella hispida Dispor eli a sp.
Escharella variolosa Escharina vulgaris Escharoides coccinea Exidmonea coerulea Fenestrulina malusii Figularia figularis Gregarinidra gregaria Hagiosynodos kirchenpaueri Haplopoma impressum Hippaliosina depressa Hippodiplosia foliacea Horner a sp.
Idmidronea atlantica Margaretta cereoides Metroperiella lepralioides Microporella appendicitiata Microporella ciliata Mimosella sp.
Mollia circumcincta
Mollia patellaria Myriapora truncata Parasmittina tropica Patinella radiata Plagioecia sp.
Puellina hincksi Puellina innominata Puellina radiata Puellina setosa Reptadeonella violacea Reteporella sp.
Rhynchozoon sp.
Savignyella lafontii Schizobrachiella sanguinea Schizomavella auriculata Schizomavella cornuta Schizomavella discoidea Schizomavella linearis Schizomavella mamillata Schizoporella magnifica Schizotheca fissa Schizotheca serratimargo Scrupocellaria delilii Scrupocellaria maderensis Scrupocellaria scrupea Sertella beaniana Smittina cervicornis Smittoidea reticulata Stephanollona armata Synnotum aegyptiacum Tubulipora liliacea Tubulipora sp. Turbicellepora sp.
Echinodermata (27 svojte) Anseropoda placenta Antedon mediterranea Arbacia lixula Astropecten irregularis Centrostephanus longispinus Cidaris cidaris Cose ‘master i as tenui spina Echinaster sepositus Echinus acutus Echinus melo Eostichopus regalis Hacelia attenuata Holothuria forskali Holothuria sanctori Holothuria tubulosa Luidia ciliaris Marthasterias glacialis Ocnus planci
Ophìacantha setosa Ophidiaster ophidianus Ophiothrìx fragilis Ophiura ophiura Ophioderma longicaudum Paracentrotus lividus Peltaster placenta Spatangus purpureus Sphaerechinus granularis
Tunicata (9 svojti) Aplìdìum conicum Aplìdìum elegans Ascidia mentula Ascidia virginea Clavelina lepadiformìs Dìazona violacea Halocynthia papillosa Microcosmus sp.
Phallusìa mammìlata
Prilog 3: FITOPLANKTON
Istraživaje fitoplanktona obavlja se na postaji Stončica od 1959. godine. Inventarna lista fitoplak-tonskih svojti načnjena je na osnovi neobjavljenih podataka Laboratorija za plankton Instituta za oceanografiju i ribarstvo, te objavljenih znanstveni radova od 1959 do 2006. godine. Popis sadrži 347 svojte (Tablica 1, Slika 1).
Slika 1. Zastupljenost pojedinih skupina fitoplanktona na postaji Stončica.
Literatura:
PUCHER-PETKOVIĆ, T., 1966: Vegetation des diatomées pélagiques de l’Adriatique moyenne. ActaAdriat. 13, 1-98.
PUCHER-PETKOVIĆ, T., MARASOVIĆ, L, 1980: Développment des populations phytoplanc-toniques caractéristiques pour un milieu eutrophisé (Baie de Kaštela, Adriatique centrale). Acta Adriat. 21, 79-93.
PUCHER-PETKOVIĆ, T., MARASOVIĆ, L, 1982: Quelques characteristiques du phytoplanc-ton dans les eaux du large de l’Adriatique centrale. Acta Adriat. 23, 61-74.
PUCHER-PETKOVIĆ, T., MARASOVIĆ, L, 1987: Contribution a la connaissance d’une poussée extraordinaire d’algues unicellulaires (Adriatique septentrionale). Centro 1, 33-44.
MARASOVIĆ, L, PUCHER-PETKOVIĆ, T., 1992: Eutrophication impact on the species composition in a natural phytoplankton community. Acta Adriat. 32, 719-72
Tablica 1. Tablični pregled zabilježenih fitoplanktonskih svojti na postaji Stončica od 1959 do 2006. godine. (*) označava svojte čiji taksonomski položaj nije sa sigurnošću određen.
ODJEL: CHRYSOPHYTA Razred: Bacillariophyceae (Diatomeae)
Achnantes sp.
Actinocvclus ehrenbergi Ralfs Actinoptychas adriaticus Grunow Actinoptychus sp.
Amphora eimotia Cleve Amphora sp. Ehrenberg ex Kützing Asterionellopsis glacialis (Castraceae) Round (=Asterionella japonica Cleve)
Asterionella bleakeleyi Smith
Astermphalus flabellatus (Brébisson) Grewille
Asterolampra marylandica Ehrenberg
Bacteriastrum delicatuhim Cleve
Bacteriastrum hyalinum Lauder
Bacteriastnm hyalinum varprinceps (Castracane)
Ikari
Bacteriastrum sp.
Biddulphia mobiliensis (Bailey) Grunow
Centricae indeterm
Cerataulina pelagica (Cleve) Hendey
Chaetoceros atlanticus Cleve
Chaetoceros anastomosons Grunow
Chaetoveros affinis Lauder
Chaetoceros affinis var. circinalis (Meunier)
Hustedt
Chaetoceros brevis Schütt
Chaetoceros circinalis (Meunier) Jensen et
Moestrup
Chaetoceros compressus Lauder Chaetoceros concavicornis Mangin Chaetoceros costatus Pavillard Chaetoceros curvisetus Cleve Chaetoceros danicus Cleve Chaetoceros diversus Cleve Chaetoceros decipiens Cleve Chaetoceros dadayi Pavillard Chaetoceros gracile Schütt
Chaetoceros laciniosus Schutt Chaetoceros lorenzianus Grunow Chaetoceros messanensis Castracane Chaetoceros perpusillus Cleve Chaetoceros pseudocrinitus Ostenfeld Chaetoceros peruvianus Brightwell Chaetoceros rostratus Lauder Chaetoceros teres Cleve Chaetoceros tetrastichon Cleve Chaetoceros tortissimus Gran Chaetoceros vixvisibilis Schiller Chaetoceros wighami Brightwell Chaetoceros sp.
ODJEL: CHRYSOPHYTA Razred: Bacillariophyceae (Diatomeae)
Achnantes sp.
Actinocyclus ehrenbergi Ralfs Actinoptychus adriaticus Grunow Actinoptychus sp.
Amphora eunotia Cleve Amphora sp. Ehrenberg ex Kützing Asterionellopsis glacialis (Castraceae) Round (=Asterionella japonica Cleve)
Asterionella bleakeleyi Smith
Astermphalusflabellatus (Brébisson) Grewille
Asterolampra marylandica Ehrenberg
Bacteriastrum delicatulum Cleve
Bacteriastrum hyalinum Lauder
Bacteriastrum hyalinum var princeps (Castracane)
Ikari
Bacteriastrum sp.
Biddulphia mobiliensis (Bailey) Grunow Centricae indeterm Cerataulina pelagica (Cleve) Hendey Chaetoceros atlanticus Cleve Chaetoceros anastomosons Grunow Chaetoveros affinis Lauder Chaetoceros affinis van circinalis (Meunier) Hustedt
Chaetoceros brevis Schütt
Chaetoceros circinalis (Meunier) Jensen et
Moestrup
Chaetoceros compressus Lauder Chaetoceros concavicornis Mangin Chaetoceros costatus Pavillard Chaetoceros curvisetus Cleve Chaetoceros danicus Cleve Chaetoceros diversus Cleve Chaetoceros decipiens Cleve Chaetoceros dadayi Pavillard Chaetoceros gracile Schütt Chaetoceros laciniosus Schutt
Chaetoceros lorenzianus Grunow Chaetoceros messanensis Castracane Chaetoceros perpusillus Cleve Chaetoceros pseudocrinitus Ostenfeld Chaetoceros peruvianus Brightwell Chaetoceros rostratus Lauder Chaetoceros teres Cleve Chaetoceros tetrastichon Cleve Chaetoceros tortissimus Gran Chaetoceros vixvisibilis Schiller Chaetoceros wighami Brightwell Chaetoceros sp.
Cocconeis scutelum Ehrenberg Coc coneis sp.
Coscinodiscus excentrions Ehrenberg Coscìnodìscus lineatus Ehrenberg Coscinodiscus sp.
Cyclotella sp.
Dactyliosolen mediterraneus Peragallo Detonui a pumila (Castracane) Gran Diploneis sp.
Diploneis bombus Ehrenberg Diploneis crabo Ehrenberg Diploneis smithii (Brébisson) Cleve Guinardia blavyana Peragallo Guinardia flaccida (Castracane) Peragallo Grammatophora marina (Lyngb.) Kützing Grammatophora sp.
Hemiaulus hauckii Grunow
Hemiaulus sinensis Greville
Eucampia cornuta Ehrenberg
Fragillaria pacifica van trigona Brun et Heribaud
Leptocylindrus adriaticus Schröder
Leptocylindrus danicus Cleve
Leptocylindrus minimus Gran
Licmophoraflabellata (Carmichael) Agardh
Licmophora grandis (Kützing) Grunow
Licmophora sp.
Melos ir a sp.
Melosira moniliformis (Müller) Agardh Melosira sulcata (Ehrenberg) Kutzing Navicula distans (Smith) Ralfs Navicula praetexta Ehrenberg Nitzchia bilobata Smith Navicula sp.
Nitzchia closterium (Ehrenberg) Smith
Nitzchia delicatissima Cleve
Nitzchia distans Gregory
Nitzchia longissima (Brébisson) Ralfs
Nitzschia paradoxa (Gmelin) Grunow (=Bacilaria
paradoxa Gmelin, B.paxillifer (Müll.) Hendy)
Nitzschia sigma (Kützing) Smith
Nitzchia tenuirostris*
Nitzchia sp.
Pennatae indeterm
Pleurosigma angulatum (Quekett) Smith Pleurosigma elongatum Smith Pleurosigma sp.
Podosira hormoides (Mont.) Kützing Pseudonitzschia spp. Peragallo Pseudo-nitzschia seriata (Cleve) Peragallo (=Nitzchia seriata Cleve)
Pseudo-nitzschia pseudodelicatissima Hasle (‘-Nitzschia delicatula Hasle )
Proboscia alata (Brightwell) Sundström Proboscia indica (Peragallo) Hemandez-Becerril Rhizosolenia calcar-avis (Schultze)
Rhizosolenia delicatula (Cleve)
Rhizosolenia fragilissima Bergon Rhizosolenia hebetata Bailey Rhizosolenia imbricata Brightwell Rhizosolenia longiseta Zacharias Rhizosolenia stolterfothii Peragallo Rhizosolenia styliformis Brightwell Rhizosolenia sp.
Skeletonema costatum (Greville) Cleve Striatella interrupta (Ehrenb.) Cleve Striatella unipunctata (Lyngb.) Agardh Sur iella sp.
Synedra pulcherrina Hantzch Synedra sp.
Thalassiosira decipiens (Grunow) Jörgensen Thalassiosira nana Lohmonn Thalassiosira sp.
Thalassionama nitzschioides Grunow Thalassiothrix frauenfeldii (Grunow) Cleve et Moller
Thalassiothrix longissima Cleve et Grunow Thalassiothrix mediterranea Pavillard
ODJEL: CHRYSOPHYTA
Razred: Chrysophyceae (Silicoflagelatae)
Dictyocha fibula Ehrenberg Distephanus crux (Ehrenberg) Haeckel Distephanus speculum (Ehrenberg) Haeckel Octactis octonaria (Ehrenberg) Hovasse
ODJEL: DINOPHYTA (Dinoflagelatae)
Amphidinium acutissimum Schiller Amphidinium britannicum (Herdman) Lebour Amphidinium conus Schiller Amphidinium crassum Lohmann Amphidinium curvatum Schiller Amphidinium flagellans Schiller Amphidinium globosum Schiller Amphidinium kesslitzi Schiller
Amphidinium klebsi Kofoid et Swetzy Amphidinium lanceolatum Schröder Amphidinium lissae Schiller Amphidinium longum Lohmann Amphidinium pelagicum Lebour Amphidinium schroderi Schiller Amphidinium stigmatum Schiller Amphidinium sp.
Ceratium arietinum Cleve
Ceratium carriense Gourret
Ceratium extensum (Gourret) Cleve
Ceratium fusus (Ehrenberg) Dujardin
Ceratium fusus var. seta (Ehrenberg) Soumia
Ceratium furca (Ehrenb.) Claparède et Lachmann
Ceratium longirostrum Gourret
Ceratium macroceros (Ehrenb.) VanHöffen
Ceratium massiliense (Gourret) Jörgensen
Ceratium palmatum Schröder
Ceratium pentagonum Gourret
Ceratium pulchelum Schröder
Ceratium reticulatum Kofoid
Ceratium symmetricum Pavillard
Ceratium teres Kofoid
Ceratium tripos (Müller) Nitzsch
Ceratium trichoceros (Ehrenberg) Kofoid
Ceratocorys armata (Schütt) Kofoid
Cladophyxis bacillifera Schiller
(=Micracanthodinium bacilliferum)
Cladophyxis brachiolata Stein Cladophyxis hemibrachiolata Balech Cladophyxis sp. Stein Cochlodinium helicoides Lebour Cochlodinium sp.
Dinophysis sacculus Stein Dinophysis sp.
Glenodinium sp.
Goniodoma polyedricum (Pouchet) Jörgensen Gonyaulax fragilis (Schütt) Kofoid Gonyaulax polygramma Stein Gonyaulax monocantha Pavillard Gonyaulax spinifera (Claparède et Lochmann) Diesing Gonyaulax sp.
Gymnodinium abbreviatum Kofoid et Swezy Gymnodinium agiliforme Schiller Gymnodinium biconicum Schiller Gymnodinium caput Schiller Gymnodinium conicum Kofoid et Swezy Gymnodinium corii Schiller Gymnodinium diploconus Schütt Gymnodinium fusus Schütt Gymnodinium gibbera Schiller Gymnodinium grammaticum (Pouchet) Kofoid et
Swezy
Gymnodinium hetero striatum Kofoid et Swezy Gymnodinium minor Lebour Gymnodinium najadeum Schiller Gymnodinium ostenfeldii Schiller Gymnodinium paulseni Schiller Gymnodinium pulchrum Schiller Gymnodinium rhomboïdes Schütt Gymnodinium simplex (Lohm) Peters Gymnodinium semidivisum Schiller Gymnodinium schroederi*
Gymnodinium simplicius *
Gymnodinium uberimum *
Gymnodinium vouki Schiller Gymnodinium sp.
Gyrodinium adriaticum Schiller Gyrodinium conicum Schiller Gyrodinium crassum (Pouchet) Kofoid et Swezy Gyrodinium falcatum Kofoid et Swezy Gyrodinium fusiforme Kofoid et Swezy Gyrodinium lachryma (Meun.) Kofoid et Swezy Gyrodinium obtusum (Schütt) Kofoid et Swezy Gyrodinium ochraceum Kofoid et Swezy Gyrodinium opimum (Schütt) Lebour Gyrodinium ovatum (Gourr) Kofoid et Swezy Gyrodinium ovum Schütt Gyrodinium pingue (Schütt) Kofoid et Swezy Gyrodinium sp.
Heterocapsa niei Murril et Loeblich (=Cachonina niei Loeblich)
Heterodinium schilleri Pavillard Massartia glauca (Lebour.) Schiller Massartia rotundata (Lohmann) Schiller Massartia sp.
Lingulodinium polyedrum (Stein) Dodge (=Gonyaulax polyedra Stein)
Oxytoxum adriaticum Schiller
Oxytoxum caudatum Schiller
Oxytoxum gladiolus Stein
Oxytoxum laticeps Schiller
Oxytoxum ovale Schiller (=0. mediterraneaum
Schiller)
Oxytoxum parvum Schiller Oxytoxum scolopax Stein Oxytoxum sphaeroideum Stein Oxytoxum viride Schiller Oxytoxum sp.
Phalacroma rotundatum (Claparède et Lach.) Kof. et Michener
Protoperidinium bipes (Paulsen) Balech (=Peridinium minusculum Pavilard) Protoperidinium brochi (Kofoid et Swezy) Balech Protoperidinium globulus (Stein) Balech
Protoperidinium granii (Ostenfeld) Balech Protoperidinium oceanicum (VanHöffen) Balech Protoperidinium steinii (Jörgensen) Balech Protoperidinium tuba (Kofoid) Balech Peridinium sp.
Pronoctiluca spinifera Lohmann Pronoctiluca sp.
Prorocentrum balticum (Lohm.) Loeblich (=Exuviella baltica Lochmann)
Prorocentrum compressum (Bailey) Abé ex Dodge (=E. compressa Stein)
Prorocentrum gracile Schütt Prorocentrum lima (Ehrenberg) Dodge (=E. marina Cienowski)
Prorocentrum micans Ehrenberg Prorocentrum scutellum Gourret Prorocentrum triestinum Schiller Prorocentrum sp.
Protodinium simplicius Schiller Phalacroma sp.
Phalacromaparvulum (Schütt) Jörgensen Podolampas bipes Stein Podolampas palmipes Stein Podolampas spinifera Okamura Podolampas sp.
Polykrikos sp.
Pyrocystis lunula Schütt Scrippsiella trochoidea (Stein) Loeblich Warnowia sp. Lindemann
ODJEL: HAPTOPHYTA Razred: Prymnesiophyceae (Coccolithophoridae)
Coccolithophoridae spp.
Acanthoica acantifera Lohmann Acanthoica coronata Lohmann Acanthoica achanthos (Schiller) Deflandre Acanthoica monospina Schiller Acanthoica quattrospina Lohmann Acanthoica sp.
Algirosphaera robusta (Lohmann) Norris (fSyracosphaera robusta Lohmann)
Anthosphaera fragaria Kamptner Armosphaera sp.
Calciosolenia granii Schiller Calciosolenia murrayii Gran Calyptrosphaera adriatica*
Calyptrosphaera dalmatica Schiller Calyptrosphaera oblonga Lochmann Calyptrosphaera globosa Lochmann Calyptrosphaera miriabilis Schiller Calyptrosphaera livella Schiller Calyptrosphaera incisa Schiller
Calyptrosphaera insignis Schiller Calyptrosphaera quadridentata Schiller Calyptrosphaera pirns Kamptner Calyptrosphaerapyriformis Schiller Calyptrosphaera sphaeroidea Schiller Calyptrosphaera tholifera Kamptoner Calyptrosphaera sp.
Coccolithus čarteri (Wallich) Kamptner Coccolithus pelagicus (Wallich) Lohmann Corisphaera arethusae Kamptner Corisphaera spinosa Kamptner Corisphaera sp.
Discosphaera thomsonii Ostenfeld Discospharea tubifer (Murr.et Black.) Ostenfeld Discosphaera sp.
Emiliana huxley (Lohmann) Hay et Mohler (=Coccolithus huxley Wall. Kampt.)
Halopappus adriaticus Schiller Lohmanosphaera adriatica Schiller Lohmanosphaera sp.
Helladosphaera cornifera (Schiller) Kamptner (=Syracosphaera cornifera Schiller) Helladosphaera sp.
Meningosphaera sp.
Ophiaster hidroideus (Lohmann) Lohmann Pontosphaera sp.
Pontosphaera echinophora Schiller Pontosphaera haeckeli Lohmann Pontosphaera nigra Schiller Pontosphaera syracusana Lohmann Rhabdosphaera hispida Lohmann Rhabdosphaera longi styl is Schiller Rhabdosphaera stylifer Lohmann Rhabdosphaera tignifer Schiller Rhandosphaera tubulosa Schiller Rhabdosphaera sp.
Rhabdonema adriaticum Kiitzing Syracosphaera adriatica Schiller Syracosphaera apsteinii Lohmann Syracosphaera brandii Schiller Syracosphaera cupulifera Schiller Syracosphaera cordiformis Schiller Syracosphaera corii Schiller Syracosphaera cornus Kamptner Syracosphaera coronata Schiller Syracosphaera dalmatica Kamptner Syracosphaera dentata Lohmann Syracosphaera grandii Schiller Syracosphaera mediterranea Lohmann Syracosphaera molischi Schiller Syracosphaera pulchra Lohmann Syracosphaera quadricornifera *
Syracosphaera quadricorni *
Syracosphaera quadridentata * Syracosphaera schilleri Kamptner Syracosphaera subsalsa (Conr.) Kamptner Syracosphaera sp.
Zygosphaera sp. Kamptner
ODJEL: EUGLENOPHYTA Razred: Euglenophyceae
Eutreptiella pascheri Schiller Eutreptia lamowii Steuer Eu trepti a sp.
ODJEL: CRYPTOPHYTA Razred: Chryptophyceae
Crypto monas sp.
Hillea fusiformis Schiller Hillea marina Butcher
ODJEL: CHLOROPHYTA Razred: Chlorophyceae
Carteria acuta Schiller Carteria crassifilis Schiller Carteria cylindracea Schiller Carteria obliqua Diesing Carteria willei Schiller Carteria sp. Diesing Chlamydomonas fusiformis Schiller Chlamydomonas minima Schiller Chlamidomonas pyriformis Schiller Chlamydomonas triangularis Schiller Razred: Prasinophyceae Pyramimonas adriaticus Schiller Tetraselmis sp. Stein
Prilog 4: ZOOPLANKON
Poj am zooplankton u moru označava sve životinj ske organizme čije su lokomotome sposobnosti nedovoljne da se opiru gibanju morskim strujama. Obuhvaća pripadnike mnoštva životinjskih skupina (protozoa i metazoa) koji se razlikuju po veličini (nekoliko pm do nekoliko m), načinu ishrane (herbivomi, detritivomi, omnivomi ili kamivomi), te periodu koji provode u planktonu. Gotovo sve veće skupine beskralježnjaka imaju svoje predstavnike u planktonu, a jaja i ličinke većine riba također se nalaze u planktonu. Od izuzetne su važnosti u hranidbenoj mreži mora, a povezani su kompleksnim trofičkim odnosima sa primamim proizvođačima – fitoplanktonskim algama kao i potrošačima koji uključuju i mnoge morske ribe i sisavce.
Podaci o zooplanktonskoj zajednici u akvatoriju otoka Visa postoje za dva lokaliteta: područje u kanalu između otoka Visa i Biševa, u neposrednoj blizini izobate od 100 m, i područje nazvano postaja Stončica (43° 00’ N; 16° 20’ E) koje se nalazi 4 Nm jugoistočno od istoimenog rta na otoku Visu.
Za područje između otoka Visa i Biševa podatke o zooplanktonu je objavio Tomo Gamulin (Gamulin, 1979) na temelju sastava svih zabilježenih skupina zooplanktona. Između ostalog, ovaj autor navodi 57 vrsta kopepodnih račića koji su među najbrojnijim višestaničnim životionjama u zooplanktonu, te povezuje brojnost i sastav kopepodne zajednice sa migracijama srdele kojoj su glavna hrana (Gamulin, 1979).
Sustavna mjesečna istraživanja zooplanktona na postaji Stončica Institut za oceanografiju i ribarstvo u Splitu započeo je još 1959. godine. Uzorkovalo se vertikalnim potegom od dna do površine pomoću Hensen planktonske mreže promjera oka svile 333 pm, a istom metodikom nastavljena su do 1991. godine.
Dubina postaje Stončica je 107 m, dnojepjeskovito ili fino detritusno sa manjim udjelom mulja. Ovo je područje otvorenog mora, visoke prozirnosti sa smanjenom brojnosti fito- i zooplanktona u odnosu na produktivnija obalna područja, ali zajednicu karakterizira visoka biološka raznolikost. Dugoročni podaci o satavu zooplanktona sa ove postaje ishodište su za više objavljenih radova s područja zooplanktona (Vučetić, 1961, 1971; Regner, 1981, 1985, 1991; Šolić i sur., 1997). Na temelju njih, Regner (1985) navodi prisustvo 90 vrsta kopepodnih račića kao najbrojnij a komponenta mrežnog zooplanktona. Osim karakterističnih vrsta kopepoda otvorenog mora, u površinskom sloju bilježi i veći broj obalnih vrsta koje se iz sjevernog Jadrana površinskom strujanjima tijekom ljeta rasprostiru do srednjeg Jadrana, ili tamo dospijevaju pojačanim strujanjem obalnih voda prema otvorenom moru.
Od 1998. godine u okviru više projekata Instituta provode se mjesečna i sezonska uzorkovanja i mrežama manjeg promjera oka (53 i 125 pm), koje daju potpunije podatke o sastavu zooplanktona na ovoj postaji jer učinkovito sakupljaju i manje zooplanktonte koje propušta mreža većeg promjera oka (Vidjak, 2007). U Tablici 1 naveden je popis 100 vrsta kopepodnih rakova koji su tijekom dosadašnjig istraživanja zooplanktona zabilježenih u viškom akvatoriju.
Od ostalih skupina zooplanktona u akvatoriju otoka Visa detaljno su analizirane i skupine želatinoznog zooplanktona (Cnidaria i Chaetognatha), a podaci o njima dostupni su u literaturi (Gamulin, 1948, 1979; Batistić, 1997).
LITERATURA:
Batistić, M. (1997) The seasonal and horizontal distribution of the biomass of epiplanktonic chaetognaths in the Adriatic Sea. Periodicum Biologorum, 99 (2): 247-254.
Gamulin, T. (1948) Prilogpoznavanjuzooplanktiona srednjedalmatinskog otočnog područja. Acta Adriat.,3 (7): 160-194.
Gamulin, T. (1979) Zooplankton istočne obale Jadranskog mora. Prirodoslovna istraživanja: Acta Biol., Vlll/l-10;270pp.
Regner, D. (1981) The changes in seasonal oscillations of copepods in the central Adriatic. Rapp. Comm. int. Mer Médit., 27 (7): 177-179.
Regner, D. (1985) Seasonal and multiannual dynamics of copepods in the middle Adriatic. Acta Adriat., 26 (2): 11-99.
Regner, D. (1991) Long-term investigations of copepods (zooplankton) in the coastal waters of the easterm Middle Adriatic. Acta Adriat. 32 (2): 631-740
Šolić M., Krstulović N., Marasović I., Baranović A., Pucher-Petković T., Vučetić T. (1997) Analysis of time series of planktonic communities in the Adriatic Sea: distinguishing between natural and man-induced changes. Oceanologica Acta 20 (1): 131-143.
Vidjak, O. (2007) Mrežni zooplankton. Kontrola kakvoće obalnog mora. Projekt Pag-Konavle. Institut za oceanografiju i ribarstvo. Split. 105-126.
Vučetić, T. (1961) Some new data on the zooplankton standing crop measurements in the Adriatic. Bilješke-Notes, 16:1-7.
Vučetić, T. ( 1971 ) Long-term zooplankton standing-crop fluctuations in the central adriatic coastal region. Thalassia Jugoslavica, 7 (1): 419-428.
Tablica 1. Popis svojti kopepodnih rakova zabilježenih na području postaje Stončica (ukupno 100 svojti).
Acartia clausi Giesbrecht, 1889 Acartia danae Giesbrecht, 1889 Acartia longiremis (Lilljeborg, 1853)
Acartia négligeas Dana, 1849 Aetidens armatus (Boeck, 1873)
Agetusflaccus (Giesbrecht, 1891)
Agetus limbatus (Brady, 1883)
Agetus typicus Kroyer, 1849 Calanus helgolandicus (Claus, 1863)
Calocalanus contractus Farran, 1936 Calocalanus neptunus Schmeleva, 1965 Calocalaniis pavo (Dana, 1849)
Calocalanus pavoninus Farran, 1936 Calocalaniis plumatus Schmeleva, 1965 Calocalanus plumulosus (Claus, 1863)
Calocalanus styliremis Giesbrecht, 1888 Candacia aethiopica (Dana, 1849)
Candacia elongata (Boeck, 1873)
Candacia giesbrecht i Grice &Lawson, 1977 Candacia tenuimana (Giesbrecht, 1889)
Candacia varicans (Giesbrecht, 1882)
Centropages kroyeri Giesbrecht, 1892 Centropages typicus Kroyer, 1849 Centropages violaceus (Claus, 1863)
Clausocalanus arcuicornis(Dana, 1849) Clausocalanusfurcatus (Brady, 1883)
Clausocalanus jobei Frost & Fleminger, 1968 Clausocalanus lividus Frost & Fleminger, 1968 Clausocalanus mastigophorus (Claus, 1863) Clausocalanus parapergens Frost & Fleminger, 1968 Clausocalanus paululus Farran, 1926 Clausocalanus pergens Farran, 1926 Copilia quadrata Dana, 1842 Corycaeus clausi F. Dahl, 1894 Ctenocalanus vanus Frost & Fleminger, 1968 Diaixis pygmoea (T. Scott, 1899)
Ditrichocoiycaeus angiicus Ditrichocorycaeus brehmi (Steuer, 1910) Ditrichocoiycaeus minimus Indicus!M. Dahl, 1912) Euaetideus giesbrechti Cleve, 1904 Eucalanus attenuatus!Dana, 1849)
Eucalanus elongates (Dana, 1849)
Euchaeta acuta Giesbrecht, 1892 Euchaeta hebes Giesbrecht, 1888 Euchaeta marina (Prestandrea, 1833)
Euchaeta spinosa Giesbrecht, 1892 Euterpina acutifrons (Dana, 1847)
Farranula rostrata (Claus, 1863)
Goniopsyllus clausi Huys & Conroy-Dalton, 2000 Haloptilus longicornis (Claus, 1863)
Heterorhabdus papiliiger (Claus, 1863)
Heterorhabdus spinifrons (Claus, 1863)
Labidocera wolastoni (Lubbock, 1856)
Lubbockia squlimana Claus, 1863 Lucicutia clausi (Giesbrecht, 1889)
Lucicutia flavicomis! Claus, 1863)
Lucicutia ovalis (Giesbrecht, 1889)
Macrosetella gracilis (Dana, 1847)
Mecynocera clausi J. C. Thompson, 1888 Mesocalanus tenuicornis (Dana, 1849)
Microsetella norvegica (Boeck, 1865)
Monothula subtilis (Giesbrecht, 1892)
Nannocalanus minor (Claus, 1863)
Neocalanus gracilis (Dana, 1849)
Oithona atlantica Farran, 1908 Oithona nana Giesbrecht, 1892 Oithona piumifera Baird, 1843 Oithona setigera (Dana, 1849)
Oithona similis Claus, 1866 Oncaea media Giesbrecht, 1891 Oncaea mediterranea (Claus, 1863)
Oncaea scottodicarloi Heron & Bradford-Grieve, 1995 Oncaea waldemari Bersano & Boxshall, 1994 Oncaea zernovi Schmeleva, 1969 Onychocorycaeus giesbrechti (F. Dahl, 1894) Onychocorycaeus ovalis (Claus, 1863)
Paracalanus denudatus Sewell, 1929 Paracalanus nanus Sars, 1907 Paracalanus pannis (Claus, 1863)
Paracandacia bispinosa (Claus, 1863)
Paracandacia simplex (Giesbrecht, 1889)
Paroithona parvula Farran, 1908 Pleuromamma abdominalis (Lubbock, 1856) Pleuromamma gracilis (Claus, 1863)
Pontella mediterranea!Claus, 1863)
Pontellopsis regalis (Dana, 1849)
Rhincalanus nasutus Giesbrecht, 1888 Sapphirina angusta Dana, 1849 Sapphirina gemma Dana, 1849 Sapphirina lactens Giesbrecht, 1892 Sapphirina nigromaculata Claus, 1863 Scolecithricella dentata (Giesbrecht, 1892) Scolecithricella tenuiserrata Giesbrecht, 1892 Scolecithrix brady Giesbrecht, 1888 Temora stylifera (Dana, 1849)
Tricoma conifera (Giesbrecht, 1891)
Tricoma dentipes (Giesbrecht, 1891)
Tricoma similis (Sars, 1918)
Urocorycaeus furcifer (Claus, 1863)
Vettoria granulosa!Giesbrecht, 1891)
Prilog 5: KITOVI
Skupina kitova (Cetacea) te dobri (Tursiops truncatus) i obični dupin (Delphinus delphis) zaštićeni su Zakonom o zaštiti prirode RH (2003) od 1995. godine. Obje vrste nalaze se u Crvenoj knjizi sisavaca Hrvatske (2006) te imaju status Ugrožen (EN) i nedovoljno poznat (DD).Također, obje vrste nalaze se na Dodatku 11 i IV Direktive o staništima (92/43/EEC).
Prema preliminarnom izvještaju o istraživanju dupina (Wiemann et al. 2007; Holcer pers.comm.) provedenom u periodu 2007-2008 utvrđeno je da u širem akvatoriju otoka Visa boravi populacija dobrih dupina. Temeljem podataka prikupljenih tijekom 3 mjeseca terenskog rada 2007. godine veličina populacije procijenjena je upotrebom Chapmanovog procjenitelja na 305 jedinki uz donji 95% interval pouzdanosti od 107 jedinki te gornji 95% interval pouzdanosti od 166 jedinki (Wiemann et al. 2007). Omjer susretanja dupina bio je 0,113 dupina/km, a učestalost opažanja 0,025 opažanja/km u uvjetima pozitivnog istraživačkog napora, dok su omjer susretanja od 0,136 dupina/ km i učestalost opažanja od 0,03 opažanja/km procijenjeni uzimajući u obzir sve vremenske uvjete te pozitivni i negativni istraživački napor (Wiemann et al. 2007).
Podaci prikupljeni tijekom 2008. još nisu obrađeni te procjena stoga još nije dopunjena ali rezultati upućuju na rezidentnost dobrih dupina unutar područja s obzirom da je tijekom dvije godine utvrđeno veće pojavljivanje istih jedinki.
Osim dobrih dupina unutar područja opažene su i druge vrste iz skupine kitova: veliki kit (Balaenoptera phvsalus), prugasti dupin (Stenella coeruleoalba) te glavati dupin (Grampus griseus) (Tablica 1).

Veliki interval odstupanja u procjeni brojnosti pokazuje da veličina i status populacije temeljem kratkotrajnog istraživanja ne mogu biti točnije utvrđeni te je stoga potrebno daljnje istraživanje. Također, potrebno je utvrditi više detalja o ukupnoj veličini područja koje navedena populacija koristi. Podaci o opažanju drugih vrsta ili pronalasku uginulih jedinki iz skupine kitova u području, dodatno naglašavaju potrebu daljnjih istraživanja ove skupine.
Literatura:
Crvena knjiga sisavaca Hrvatske. Nikola Tvrtković (urednik). 127 str. Ministarstvo kulture Republike Hrvatske : Državni zavod za zaštitu prirode, 2006.
EU Habitats Directives (1992) The Council Of The European Communities Directive 92/43/EEC on the Conservation of Natural Habitats and of Wild Fauna and Flora
Zakon o zaštiti prirode (2003) Narodne novine 162.
Wiemann A, Holcer D, Fortuna CM & Mackelworth PC (2007): Adriatic Dolphin Project Vis. Report on the establishment of a preliminary field base and the first cetacean surveys around the island of Vis. Blue World Institute of Marine Research and Conservation. Veli Lošinj. 8pg.
Tablica 1. Popis vrsta iz skupine kitova (Cetacea) zabilježenih na području Viškog arhipelaga.
dobri dupin Tursiops truncatus veliki kit Balaenoptera phvsalus prugasti dupin Stenella coeruleoalba glavati dupin Grampus griseus
Prilog 6: Izvještaj s javne prezentacije projekta
Prezentacija projekta pod nazivom „OČUVANJE I ODRŽIVO KORIŠTENJE PODMORJA VIŠKOG ARHIPELAGA“ održana je u Komiži 07.studenog 2008. u galeriji Boris Mardešić. Prezentacije je trajala tri sata i obuhvatila je prikaz rezultata istraživanja, prezentaciju prijedloga za održivi razvoj, prezentaciju promotivnog materijala napravljenog u sklopu projekta te raspravu. Na prezentaciji je sudjelovalo oko 20 profesionalnih ribara s otoka Visa, gradonačelnica grada Komiže, nekoliko ribara dopunaca te nekolicina zainteresiranih ronioca rekreativaca.

od pet promotivnih filmova (Medvidova spilja, Zelena spilja, Vasillios, Plavetnilo, Gorgonije) načinjenih u sklopu projekta kao i prezentacija info ploče o podmorju Biševa.
Prezentaciju projekta obavili su:
Dr. se. Ante Žuljević, s prikazom istraživanja bentoskih zajednica, njihovih ugroza i mogućnosti iskorištavanja;
Dip. inž. Nika Sagličić, s prikazom ihtiofaune istraživanog područja;
Dr. se. Sanja Matić-Skoko, s prikazom istraživanja priobalnog ribolova Viškog akvatorija, analizom kategorija ribara, plovila i korištenih ribolovnih alata, analizom ulova te preporukama za očuvanje ekosustava, a posebno komercijalnih resursa u smislu održivog ribolova;
Dipi. inž. VedranNikolić, sprikazom pozitivnih svjetskih primjera osnivanja zaštićenih morskih područja
Dr.se. Ante Žuljević s prijedlogom uspostave ribolovnih zona otoka Visa i dva posebno upravljana područja na otoku Biševu.
Između svake prezentacije prikazan je jedan

Prezentacije su pokazale kako na području Visa i Biševa postoji iznimna raznolikost vrsta i staništa. Ova raznolikost podržava razvoj gospodarski interesantnih vrsta riba i rakova. Međutim, preveliki ribolovni napor dovodi do smanjenja ulova komercijalno zanimljivih vrsta. Posebno je zabrinjavajući trend da se love sve manje jedinke, dok su one velikih tjelesnih dužina i masa vrlo rijetke. U cilju očuvanja i obnove ekosustava te komercijalno isplativih lovina, ali i nove, obogaćene turističke ponude, predložene su slijedeće mjere:
1. uspostava ribolovnih zona na cijelom području Visa
2. uspostava posebno upravljanih područja na Biševu.
Ribolovne zone postojale su za područje Visa i okolnih otoka (Biševo, Jabuka, Brusnik) do početka 90tih godina prošlog stoljeća. Profesionalni ribari se slažu kako je to bila pozitivna praksa koja je prestala zbog političko povijesnih razloga jer je stvaranjem Republike Hrvatske došlo do ukidanja općinskih ovlasti pod čijom je nadležnosti bila ova regulativa. Slažu se kako bi povratak zona bilo dobro rješenje, ali se boje da će najveći problem biti u slaboj kontroli zabrane kao i da se ista neće odnositi na sve kategorije ribolova.
Zaključeno je da se nastavi rad u smislu
određivanja načina i dinamike ponovne uspostave zona s dvogodišnjom rotacijom.
Uspostava posebno upravljanih područja s tzv. „no take“ zonama prihvaćena je kao izvrsna ideja. „No take“ zone pružaju mogućnost obnove ekosustava, sazrijevanja komercijalnih vrsta te njihovo „prelijevanje“ izvan takve zone.
Ovakva područja su turistički zanimljiva zbog svoje „nedirnute“ bioraznolikosti i kao takva su izvrstan turistički brend i imaju veliki potencijal iskorištavanja posebno u ronilačke svrhe. Međutim, predložene zone na otoku Biševu (prva oko Modre špilje i druga oko Medviđe špilje) smatraju se najboljim ribolovnim postajama, pa je sugerirano da se one organiziraju s ponešto izmjenjenom površinom i lokacijom (na istočnoj strani Biševa) gdje bi bio minimalan sukob interesa s ribarima. Dodatno, gradonačelnca je istaknula kako za područje Modre špilje grad ima koncesiju za dovođenje turista u obilazak što bi s uvođenjem „no take“ zona postalo neizvedivo.
Zaključeno je, da kod drugog prijedloga postoji cijeli niz otvorenih pitanja koje zahtjevaju da sve zainteresirane strane iznesu mišljenje, i da se kroz raspravu dođe do rješenja koje bi zadovoljilo sve strane, a sve u svrhu očuvanja neupitnih ljepota i bioloških vrijednosti izabranih lokaliteta.

Projekt COAST razvijen je uz potporu Programa Ujedinjenih naroda za razvoj (UNDP), u suradnji s Ministarstvom zaštite okoliša, prostornog uređenja i graditeljstva tedrugim nadležnim ministarstvima,4dalmatinske županije te brojnim lokalnim udrugama, tvrtkama i pojedincima, a provodi se uz financijsku potporu Globalnog fonda za okoliš (GEF). Planirano trajanje projekta je 7 godina. Lokalni ured za provedbu Projekta smješten je u Splitu.
Projekt obuhvaća obalno područje četiri dalmatinske županije: Zadarske, Šibensko-kninske, Splitsko-dalmatinske i Dubrovačko-neretvanske. Unutar tog područja prepoznata su i odabrana 4 demonstracijska područja zbog svoje iznimne biološke i krajobrazne vrijednosti. To su (1) Pelješac, Dubrovačko primorje, Malostonski zaljev i Mljet; (2) Vis i viški akvatorij; (3) šire područje ušća rijeke Krke; te (4) otok Pag (jugoistočni dio u Zadarskoj županiji), područje uz Novigradsko i Karinsko more.
Osnovni cilj projekta COAST je učinkovito utjecati na poduzetničke aktivnosti i prakse u turizmu, poljoprivredi, ribarstvu i marikulturi, izravno i kroz bankarski sektor, kako bi isti u svoje prakse uključili održivo korištenje i očuvanje biološke i krajobrazne raznolikosti.
Očuvanje biološke raznolikosti dalmatinske obale, kroz promicanje održivog razvoja, moguće je jedino kroz suradnju svih partnera na nacionalnoj, županijskoj i lokalnoj razini, kao i svih zainteresiranih strana, prvenstveno na području Dalmacije.
Klima otoka Biševa
Klima Viškog arhipelaga, pa tako i Biševa, blaga je i ugodna u usporedbi s klimom drugih naših krajeva, pa i Južne Hrvatske. Ako se usporede njegove temperaturne vrijednosti s užim područjem srednjodalmatinske obale, vidljivo je da je temperatura ovdje nešto niža nego na obali od ožujka do rujna, i to od jedne polovice do jednog cijelog stupnja po Celsiusovoj skali. U tijeku zime, od listopada do uključivo veljače do dva stupnja je viša nego na obali. U tome veliku ulogu igra more. Ono se ljeti teže zagrijava od kopna, a zimi polakše hladi, i tako ublažuje vrućinu ili hladnoću. Srednja temperatura na Visu za razdoblje od 1958. do 1961. iznosila je +16,7° C, s najvišom +35,6° C i najnižom -2,4° C.
Bura i sjeverni vjetrovi blaži su u Viškom arhipelagu, pa tako i na Biševu, u usporedbi s kopnom jer na putu od kopna do ovih otoka oslabe. Jugoistočnjak ili jugo kako ga narod ovdje naziva, podjednakom jačinom puše ovdje i na obali. Na ovim otocima puše 19% dana u godini. Sjeverozapadnjak, maestral, puše u toplijem dijelu godine, ljeti oko 25% dana; jači je ovdje nego na obali i vrlo ugodno osvježava vruće ljetne večeri.
Oborine su u Viškom arhipelagu, pa i na Biševu, oskudne. Od 1958. do 1961. količina oborina iznosila je 575,9 cm u prosjeku; obilnije su zimi nego ljeti. Na kraju treba istaknuti jednu zanimljivost. Za vrijeme ljetnih vrućina na otoku Visu noći su suhe. Tek pred jugo noću padne rosa. Na Biševu je obratno. Za vrijeme ljetnih vrućina svake noći pada obilna rosa. Noć pred jugo bude suha bez rose.
Na Biševu obilno raste bor /Pinus halepensis/. Na istočnom dijelu otoka je bujna borova šuma. Nekada je bila veoma gusta, ali ju je 1936. godine zahvatio veliki požar. Dobrim dijelom bila se oporavila ali ju je požar 1994. ponovo uništio. j\|. Biševu raste česvina ili kako je narod zove česmina /Quercus ilex/, lovorika /L^ rus nobilis/, zelenika /Phillyrea media/, smreka /Juniferus Oxicedrus/, vrijes /Eric, arborea/, ružmarin /Rosmarinus officinalis/ sa svojim karakterističnim cvijetom j mirisom, mrča /Myrta italica/, smrdljika /Pistacia Terebinthus/, žuka /Sparitiu^ junceum/, drača /Paliurus australis/ i kupina /Rubus ulmifolius/. Na Biševu su j busovi tipične sredozemne makije, i to trešlje, bušine, kozje krvi i kičice /Centarj. um speciale/, koja se ističe ružičastim cvijećem, i grmovi Gospine trave /Hyper icum perforatum/ sa žutim cvjetovima. Neki dijelovi kamenjara obrasli su niskim grmljem smilja karakterističnih zlatnožutih glavica s dlakavom bjeloglavicom koja cvate ružičastim cvjetovima. U pukotinama kamena raste kapara /Caparis spinosa/. Po otoku raste agava /agauos/, biljka iz porodice sunovrata /amarylliaceae/, j kupina / Rubus ulmifolius/. Od kulturnih biljaka dobro uspijeva loza /Vitis vinifera/, smokve razne vrste, od ranih do kasnih, badem /Amygdalus communis/, oskoruša /Lorbus domestica/. Na Biševu uspijeva posebna vrsta mirisne breskve koju narod naziva breskva golica. Mogao bi uspijevati rogač /Ceratonia siliqua/, a i masline /Olea europea/. Ljudi su u prošlom stoljeću sjekli masline i na njihovo mjesto sadili lozu, jer je loza bila rentabilnija. Mogao bi uspijevati limun /Citrus limonum/ i naranča /Citrus aurantinum/, ali narod ih rijetko sadi zbog štednje vode. Povrtlarstvo bi jednako moglo dobro uspijevati, ali uzgaja se samo za kućne potrebe zbog štednje vode i nepovoljne veze s tržištem. Jedino se na veliko uzgajao zeleni grašak /Pisum sativum/. Voće i povrće na Biševu sazrijeva 52 dana ranije nego u Podunavlju.
Na Biševu raste uskolisna zelenika /Phillyrea media/. Vrlo rijedak endem koji se nalazi još jedino u okolini Dubrovnika. Na padinama luke Mezuporat nalazi se zanimljiva biljka, neka vrst čupavca /Mesembriantemum acinacioforme/ koji je prenijet iz Egipta i ovdje se udomaćio.
Podmorska vegetacija oko Biševa spada u podmorsku vegetaciju takozvanog vanjskog pojasa jer je karakterizira niz biljnih vrsta kojih nema u vodama otoka bližih jadranskoj obali. Među njima se posebno ističu smeđe alge Cistosire koje su se razvile samo u Jadranu, a u drugim morima ih nema. Značajno je da podmorska vegetacija u ovim vodama doseže znatno veću dubinu nego u vodama otočja koje se nalazi uz unutarnje obalno područje. U biševskim vodama se nalaze pojedini primjerci algi na sto metara dubine. U moru su poznate sipe /Sepia officinalis/ hobotnice i lignje / loligo vulgaris/, te nekoliko vrsta morskih ježeva i zvijezda. Inače, prvo je karakteristično za vanjski dio Jadrana, a drugo za čitavo Jadransko more. Još treba spomenuti školjke kao periske /Pinna squanosa/ i neke vrste oštriga. Sve do nedavno pojave velikih jata plave ribe u ovim vodama svjedoči o bogatstvu ovoga mora planktonima i rakovima veslonošcima, kojima se poglavito hrani plava riba. Na brakovima i podmorskim grebenima oko Biševa živi jastog /Palmirus vulgaris/. U jesen ovdje se love iglice /Geranium/.
Na otoku Biševu nema otrovnih zmija. Ovdje se nailazi na šaru poljaricu /Zamensis genomensis, Laur/, pjegavu crvenkapicu /Elapha situla, Laur/ i na našu najdužu zmiju, kravosca /Coluber flavescens/. Na Biševu živi 61 vrsta kornjača. Od njih 36 vrsta živi na čitavoj dalmatinskoj obali, 22 na srednjodalmatinskim otocima, a 3 vrste su nađene jedino na Biševu i na najudaljenijim otocima Jadranskog mora. Na ovom otoku ima 14 vrsta mrava i 7 vrsta cvrčaka.
Ni najstariji ljudi se ne sjećaju da bi se na Biševu u većem broju uzgajalo ovce ili koze, tek poneku za obiteljske potrebe. Jednako tako se držalo magarca, kobilu, mazgu ili mulu, koliko je pojedinim obiteljima bilo potrebno.
Među vodozemcima na Biševu živi zelena gubavica, a među guštericama ima jedna endemske vrste: Lacerta fiumana lissana. To je zapravo poseban oblik primorske gušterice koja živi na kopnu i na otocima u posebnim oblicima. Na ovom otoku ima i jedna vrsta bezopasnog, dapače korisnog, zidnog guštera – macaklin /Hemidactylus turcina/. Ljudi ga se boje i nazivaju gubavicom, smatraju ga otrovnim, a to on nije.
Od sisavaca na Biševu živi nekoliko vrsta sitnijih glodavaca. Od njih najveći je obični zec /Lepus europeus/. To je posebna podvrsta zeca nižeg rasta. Od sisavaca kukcojeda na otoku živi jež /Erinaceus romanicus/, a od netopira brojne vrste šišmiša.
Istraživanja i zaštita
Na Biševu se nalaze dva geomorfološka spomenika prirode: Modra spilja (zaštićena 1951. god.) i Medvidina pećina na južnoj obali otoka, azvana po svom nekadašnjem stanovniku morskoj medvjedici (Monachus monachus Herman) (zaštićena od 1967.) Područje je u sastavu Nacionalne ekološke mreže: Biševo kopno (HR2001097) i more (HR3000098), Modra špilja (HR3000184) i Medvidina pećina (HR3000205), od međunarodne je važnosti za ptice. IPA je također podložno primjeni Uredbe o uređenju i zaštiti zaštićenog obalnog područja mora (NN 128/04) (obuhvaća sve otoke, pojas kopna u širini od 1000 m od obalne crte i pojas mora u širini od 300 m od obalne crte). Na području je obavljeno kartiranje flore i staništa u okviru projekta COAST (Conservation and Sustainable Use of Biodiversity in the Dalmatian Coast through Greening Coastal Development), te IPA projekta (Important Plant Area – Croatia) 2007 i 2008 god. Također, provedena je analiza bioraznolikosti u sklopu projekta «Akcijski plan za očuvanje Dalmacije».
Staništa i svojte
Šuma crnog jasena i hrasta crnike (Fraxino orno-Quercetum ilicis), koja klimazonalno dolazi na ovom području, na otoku Biševu je razvijena u obliku degradiranog stadija – makije. Osim makije, ovdje dolaze i bušici crnjuše i ružmarina (Erico multiflorae-Rosmarinetum), kao daljnji stadij degradacije šume. Veći dio otoka zauzimaju suhi travnjaci, u obliku kamenjara raščice i zvjezdaste djeteline (Brachypodio retuso-Trifolietum stellati) te travnjaka prosuljastog šćevara (Oryzopsetum miliaceae). Za kamenite morske obale značajna je grebenjača rešetkaste mrižice i trpuca (Plantagini-Limonietum cancellati), dok se na stijenama razvija zajednica busine i dubrovačke zečine (Phagnalo-Centaureetum ragusinae). Od vegetacije koja je vezana za nazočnost čovjeka dolazi zajednica slezenice i zidnog kločućića (AsplenioUmbilicetum horizontalis) kao predstavnik vegetacije pukotina starih zidova, te nekoliko korovnih i ruderalnih zajednica: zajednica babjeg zuba i šćira (Tribulo-Amarantetum), zajednica drvolike stole (Lavateretum arboreae), zajednica primorskog divljeg ječma (Hordeetum leporini) i utrina ljulja i busenastog trpuca (Lolio-Plantaginetum commutatae). Među značajnijim tipovima vegetacije treba spomenuti sastojinu kritično ugrožene valjkaste zupčice (Imperata cylindrica) male površine (nekoliko stotina kvadratnih metara), razvijenu na pjeskovitoj podlozi na području Gornje Salbunare. Na Biševu je do sada registrirano oko 480 biljnih vrsta. Biševo je značajno kao tipično nalazište (locus classicus) za Stalijevu modričicu (Asperula staliana ssp. staliana) i pjeskovitu modričicu (Asperula staliana ssp. arenaria), kojoj je to ujedno i jedino poznato nalazište. Osim toga, otok je jedno od rijetkih nalazišta končaste mlječike (Euphorbia terracina – napuštena polja i terasasti pješčani vinogradi) i pilastog trpuca (Plantago serraria), te jedno od tri nalazišta Mihelijeve avelinije (Avellinia michelii), uz otoke Vis i Hvar. Na otoku je nekada rasla i rijetka vrsta otantus (Otanthus maritimus), međutim već dulje vrijeme nije ovdje zabilježena. U sastavu makije raste i globalno ugrožena vrsta šaša (Carex illegitima), flora pješčanih staništa je dobro razvijena, te ovdje nalazimo i rijetku vrstu primorskog kotrljana (Eryngium maritimum). Ovdje dolazi veći broj endemičnih i ugroženih biljaka, od kojih pješčana sijačica (Sporobolus pungens) nije potvrđena na otoku već dulje vrijeme.
Bilješke
Otok Biševo kao i cijeli Viški arhipelag nalazi se u eumediteranskoj zoni. Analizom flore otoka Biševa do sada je zabilježeno 485 vrsta viših biljaka (PAVLETIĆ 1974; BOGDANOVIĆ et al 2004; FCD) (Prilog 3). Ovaj, iako malen otok u biljnogeografskom pogledu veoma je zanimljiv na što su još davno ukazali mnogi botaničari (VISIANI 1842-1850; GIZBENGER 1921). Na otoku je dominantna zajednica gariga koja pripada asocijaciji Erico-Rosmarinetum iz sveza Cisto-Ericion i to pretežito na neobrañenim dijelovima, a česte su i zajednice makije iz iste sveze te zajednice kamenjarskih travnjaka iz reda Cymbopogo-Brachypodietalia, iz sveze CymbopogoBrachypodion ramosi. Na kamenjarskim travnjacima iz asocijacije Brachypodio-Trifolietum stellati i Brachypodio-Cymbopogonetum hirti i garizima oko Salbunare i Polja dolaze rijetke i ugrožene biljke poput vrste Avellinia michelii, kojoj su otoci Biševo, Vis i Hvar zasad jedini poznati lokaliteti i vrsta Heteropogon contortus sa svega par lokaliteta u središnjoj i južnoj Dalmaciji. Na južnom dijelu otoka nalazi se razvijena prirodna mediteranska vazdazelena šumska zajednica hrasta crnike, koja pripada razredu Quercetea ilicis i asocijaciji Fraxino orni-Quercetum ilicis, a na pojedinim mjestima je razvijena 2Mediteranskom flornom elementu pripada 80% flore otoka, od toga 70% pripada istočno-mediteranskom, 25% istočnomediteranskom, a 5% su ostali elementi. 3 Jadranski, centralno-mediteranski, sjeverno-mediteranski, južno-mediteranski i egejski element. 4 Pontsko-mediteranski, srednje-europski, atlanski florni element. 75 kao šuma alepskog bora (Pinus halepensis). U tim šumama dolazi jedna rijetka vrsta šaša, Carex illegitima, koja se ne nalazi u Crvenoj knjizi, ali je vrijedna pažnje. U središnjem dijelu otoka na zaravnjenoj blagoj udolini podloga je pješčana te se razvila vegetacija pijesaka, koja pripada asocijaciji Agropyretum mediterraneum sa stenoendemičnom, ugroženom, pjeskovitom modričicom Asperula staliana subsp. arenaria (kojoj je otok Biševo ujedno I jedino poznato nalazište – locus classicus) i ugroženim žiljem, Pancratium maritimum. Za pijeske su vezane i neke druge rijetke i ugrožene biljke, kao što su Sporobolus pungens, sa svega par nalazišta u Hrvatskoj i vrsta Imperata cylindrica, koja gradi sastojinu veličine nekoliko hektara u okolici Gornje Salbunare. Uz morsku obalu razvijena je hazmofitska endemična asocijacija Pagnalo-Centaureetum ragusinae. Toj zajednici, koja je razvijena u potpunosti, pripadaju neke rijetke, endemične i zakonom zaštićene biljke. To su dubrovačka zečina, Centaurea ragusina i Convolvulus cneorum. U sastavu vegetaciji stijena raste još jedna zakonom zaštićena vrsta, grmoliki (Jupiterov) ranjenik, Anthyllis barba-jovis. Halofilna vegetacija je ovdje jako dobro razvijena sa svim svojim karakterističnim vrstama i pripada razredu Crithmo-Limonietea i asocijaciji Plantagini-Limonietum cancellati u kojoj dolazi endemični Limonium cancellatum i Senecio leucanthemifolius var. reichenbachii, koja je rijetki endemični varijetet hrvatske flore. Meñu značajnijim tipovima vegetacije treba spomenuti sastojinu kritično ugrožene valjkaste zupčice (Imperata cylindrica) male površine (nekoliko stotina kvadratnih metara), razvijenu na pjeskovitoj podlozi na području Gornje Salbunare. Analizom flornih elemenata prema PAVLETIĆ (1975.) sve su biljke svrstane u tri skupine: mediteranski florni element, južnoeuropski florni element i biljke širokog rasprostranjenja. Najvažniju ulogu u izgradnji biljnog pokrova ovog otoka ima mediteranski florni element, osobito pripadnici općemediteranskih biljaka. Ta činjenica potvrñuje pripadnost Biševa (u biljnogeografskom smislu) eumediteranskoj regiji. Osim visokog postotka općemediteranskih biljaka (Tab. 10) znatna je prisutnost vrsta koje flori, a time i vegetaciji Biševa daju specifičan karakter. Prvenstveno se radi o tzv. ilirsko-mediteranskim biljkama meñu kojima osobitu ulogu imaju ilirsko-jadranske endemične biljke. S druge strane, ilirsko-apeninske biljke, uz neke vrste zapadno-mediteranskih biljaka, kojima je otok Biševo istočna granica rasprostranjenja, ukazuju na biljnogeografsku povezanost naših srednjodalmatinskih otoka i Apeninskog poluotoka. Ta veza je uvjetovana, kao što je poznato, vezom iz geološkoj prošlosti u periodu dok su ti dijelovi bili cjelovito kopno. Relativno velika zastupljenost zapadno-mediteranskih biljaka govori u prilog i tvrdnji TRINAJSTIĆA (1973.) da je upravo u području otočne skupine Visa jedno pribježište ovih biljaka u nas. Tablica 10. Međusobni odnos flornih elemenata (prema PAVLETIĆ, 1975) na otoku Biševu. I. MEDITERANSKI FLORNI ELEMENT 64.3% A. OPĆEMEDITERANSKE BILJKE 41.2% B. ZAPADNOMEDITERANSKE BILJKE 4.7% C. ISTOČNOMEDITERANSKE BILJKE 2.6% D. ILIRSKOMEDITERANSKE BILJKE 4.4% E. MEDITERANSKO-ATLANSKE BILJKE 4.7% F. EVROPSKO-MEDITERANSKE BILJKE 3.6% G. MEDITERANSKO-PONTSKE BILJKE 3.1% II. JUŽNOEVROPSKI FLORNI ELEMENT 13% A. JUŽNOEVROPSKO-MEDITERANSKE BILJKE 10.7% B. JUŽNOEVROPSKO-PONTSKE BILJKE 2.3% III. BILJKE ŠIROKOG RASPROSTRANJENJA 22.7% Usporedbom pojedinih životnih oblika biljaka otoka Visa i Biševa dolazi se i do zaključka da Biševo ima sušu klimu nego otok Vis, što upućuje i visok postotak terofita (51.4%) (Tab. 11).
Tablica 11. Usporedba životnih oblika biljaka otoka Visa i Biševa (prema PAVLETIĆ, 1975). Ph% Ch% H% G% T% VIS 10 7 26 11 46 BIŠEVO 14.4 17.7 10.8 5.7 51.4 Od posebnog su značaja za floru Biševa pa i Hrvatske, neke rijetke, ugrožene i endemične biljne svojte, kojima je Biševo jedno od rijetkih nalazišta u Hrvatskoj. 76 Istraživanjem flore i orhideja otoka Biševa 2000. i 2001. godine u okviru projekta studentske udruge “BIUS”, zabilježene su dvije endemične vrste orhideja; Ophrys x lyrata (=O. bertolonii x O. incubacea) i Ophrys sphegodes subsp. tommasinii (=O. tommasinii) te jedna ugrožena vrsta, Cephalanthera longifolia i rijetka vrsta hrvatske flore, Ophrys bombyliflora sa svega par poznatih nalazišta u Hrvatskoj (BOGDANOVIĆ, DOBROVIĆ i OSTOJIĆ, 2001). Endemična biljka otoka Biševa (prema PAVLETIĆ, 1978.) koja nije uvrštene u Crvenu knjigu biljnih vrsta Republike Hrvatske je Senecio leucanthemifolius var. reichenbachii. Ovu biljku bilježi Fiori za manje otoke kraj Sardinije te Palagružu i Tremite u Jadranu (FIORI 1923-1929). GINZBERGER 1921. godine navodi ovaj varijetet za otoke Sušac, Veliku i Malu Palagružu, a za Brusnik navodi Senecio leucanthemifolius Poir. var. pinnatifidus Fiori. Do sada poznata nalazišta su Palagruža (GINZBERGER 1921; PAVLETIĆ 1978), Sušac (GINZBERGER 1921), meñutim Korica u svojim istraživanjima ne bilježi tu vrstu (KORICA 1977), Brusnik (PAVLETIĆ 1970, 1978), Biševo (PAVLETIĆ 1974, 1975), Svetac (PAVLETIĆ 1978, 1979), Jabuka i Kamik (PAVLETIĆ 1978a). U talijanskom dijelu Jadrana dolazi i na Tremitima (prema PAVLETIĆ 1975). Ova svojta nam govori o biljnogeografskoj povezanosti ovih otoka u periodu postgalcijala dok su ovi otoci bili povezani s Apeninskim poluotokom (BOGDANOVIĆ, 2002.). U flori otoka Biševa utvrñene su 43 biljne vrste koje pripadaju različitim kategorijama ugroženih vrsta, 88 zaštićenih biljnih vrsta, 7 endemičnih biljnih vrsta, 19 biljnih vrsta koje su na popisu zaštićenih vrsta Bernske konvencije (označene u prilogu popisa oznakom BK)
Otok Vis s otocima
Klimazonalna vegetacija većeg dijela otoka Visa je “čista, vazdazelena šuma i makija crnike s mirtom” (Myrto-Quercetum ilicis) Trinajstić 1985). Ipak, najdominantniji tip staništa na otoku su „zapadnomediteranski bušici ružmarina“ (Rosmarinetalia). Ovaj tip staništa dominira većim dijelom otoka i dolazi u kombinaciji s drugim tipovima staništa od obale mora do Huma, najvišeg vrha otoka Visa. Vrlo često na zapuštenim poljoprivrednim površinama, najčešće bivšim vinogradima, dolazi u kombinaciji s daljnjim degradacijskim stadijem, a to su „eu- i stenomediteranski kamenjarski pašnjaci raščice“ (Brachypodio-Cymbopogonetum hirti H-ić. 1961). Velike površine „zapadnomediteranskih bušika ružmarina“ (Rosmarinetalia) nalaze se na prijelazu u sljedeći sukcesijski stupanj klimazonalne makije crnike ili obrastaju šumama alepskog bora.
Šume alepskog bora u južnijim dijelovima otoka većim dijelom pripadaju stanišnom tipu „šume alepskog bora sa sominom” (Junipero phoeniceae-Pinetum halepensis Trinajstić 1988), dok manjim dijelom dolaze kao “šuma alepskog bora s tršljom” (Pistacio-Pinetum halepensis De Marco, Veri et Caneva 1984). U središnjem i sjevernom dijelu otoka uglavnom prevladava „mješovita šuma alepskog bora i crnike” (Querco ilicis-Pinetum halepensis Loisel 1971). Ova podjela je samo okvirna, jer vrlo često na površinama koje su označene kao “mješovita šuma alepskog bora i crnike“ nema puno crnike, već su ovim stanišnim tipom opisane i sve one površine gariga koje su obrasle alepskim borom. Premda se fitocenološki ovakva vegetacija opisuje kao facies Pinus halepensis asocijacije EricoRosmarinetum, prilikom kartiranja staništa ovakav bi pristup bio pogrešan. Također, iz prikupljenih podataka nije bilo moguće dovoljno dobro odvojiti „šume alepskog bora sa sominom” (Junipero phoeniceae-Pinetum halepensis Trinajstić 1988) od “ šuma alepskog bora s tršljom” (Pistacio-Pinetum halepensis De Marco, Veri et Caneva 1984). Na području vrha Hum, na sjeverno eksponiranim padinama nalazimo stanišne tipove s listopadnim elementima. Na samom vrhu dolazi stanišni tip vlažnijih i hladnijih područja “mješovita šuma i makija crnike s crnim grabom” (Ostryo-Quercetum ilicis Trinajstić (1965) 1977). Na nešto nižim nadmorskim visinama, a još uvijek na sjeverno eksponiranim područjima, nalazimo veće površine „mješovite šume i makije crnike s crnim jasenom” (Fraxino orni-Quercetum ilicis H-ić. (1956) 1958). Oba stanišna tipa dolaze u obliku makije. Također, na samom području vrha Hum, na degradiranijim dijelovima makije mozaično se pojavljuju dijelovi s „istočnojadranskim kamenjarskim pašnjacima submediteranske zone” (Chrysopogoni-Koelerion splendentis H-ić. 1975), na kojima uglavnom dominiraju Salvia officinalis i Satureja montana. U ostalim dijelovima otoka Visa najčešći tip makije je makija crnike s mirtom (Myrto-Quercetum ilicis (H-ić.) Trinajstić 1985). Samo na južnom dijelu otoka, u blizini morske obale dolazi i makija tršlje i somine (Pistacio-Juniperetum phoeniceae Trinajstić 1987). Najveće travnjačke površine zauzimaju „eu- i stenomediteranski kamenjarski pašnjaci raščice“ (Brachypodio-Cymbopogonetum hirti H-ić. 1961), koji i najčešće dolaze u obliku jednoličnih sastojina u kojima dominira Brachypodium retusum. U istočnom dijelu otoka Visa na nekim se napuštenim poljima nalaze veće površine “jadranskih travnjaka brčka” (Vulpio-Lotion H-ić. 1960).
Najznačajnija poljoprivredna kultura na otoku Visu su vinogradi. Od ostalih kultura dolaze sporadično mali tradicionalni maslinici, te pokoja zapuštena poljoprivredna površina koja se kosi. Još uvijek se na otoku mogu naći rogačici, ali oni zauzimaju vrlo male i sporadično raspoređene površine. Od ostalih kultura potrebno je izdvojiti nedavno zasađen (poslije 2004. godine) veliki voćnjak na padinama koje gledaju prema uvali Mala Smokova. Na području Smokovo, područje bivše vojarne, nalazi se jedna od rijetkih površina na otoku Visu koja se još koristi kao pašnjak. Ostale poljoprivredne površine su u različitim fazama zarastanja. Ako su takve površine zarasle u bušik ili makiju, prikazane su s pripadajućim stanišnim tipom i na karti staništa. U ostalim slučajevima su zarasle poljoprivredne površine opisane stanišnim tipom “mozaika poljoprivrednih površina i prirodne vegetacije s prevladavajućom grmolikom vegetacijom i drvećem”.
Od rijetkih i ugroženih tipova staništa potrebno je istaknuti područje Velike Smokove na kojem se nalaze “površine pješčanih plaža pod halofitima” na kojima dominira Pancratium maritimum. Na više mjesta na obali otoka Visa nalaze se razvijene “dalmatinske vapnenačke stijene”, no zanimljiv je nalaz ovoga tipa staništa na stijenama u uvali Pritišćina, jer se nalazi vrlo nisko. Novoizgrađena cesta do uvale Pritišćina, a time i povećan broj turista, mogli bi utjecati na izgled ovog lokaliteta u budućnosti. Premda na otoku postoje brojne pješčane i šljunčane plaže, najveći dio je “stjenovita morska obala” na kojoj dominiraju “površine stjenovitih obala pod halofitima”.
Brusnik i Jabuka
Otoci Brusnik i Jabuka su vulkanskog porijekla, te imaju slabo razvijenu vegetaciju. Na oba otoka je razvijen endemični stanišni tip „grebenjače maljave mrižice i šćulca“ (Crithmo-Limonietum vestiti Trinajstić in Zi.Pavletić 1989). To je stanišni tip priobalnog dijela zone zračne posolice i prskanja morskih valova.
Na otoku Jabuci se u područjima iznad zone zračne posolice i prskanja morskih valova nalaze stanišni tipovi „ljeti listopadnog grmljaka drvenaste mlječike“ i „neobrasle i slabo obrasle stijene“. Veći dio otoka Brusnika je obrastao „zajednicom drvolike stole“ (Lavateretum arboreae Br.-Bl. et Molinier 1935), a u središnjem dijelu otoka na strmim stijenama nalazimo vrstu Centaurea ragusina.
Na otoku Jabuci i Brusniku dolaze biljke iz sveze Centaueo-Portenschlagiellion Trinajstić 1980, odnosno zajednice Phagnalo-Centauretum ragusinae H-ić 1962, koje su opisane stanišnim tipom „dalmatinskih vapnenačkih stijena“ (B.1.4.2.). Kako su Jabuka i Brusnik otoci vulkanskog porijekla, na kojima nema vapnenaca, nismo iz formalnih razloga na ovim otocima mogli opisati površine sveze Centaueo-Portenschlagiellion Trinajstić 1980 kao stanišni tip „dalmatinske vapnenačke stijene“. U vezi s tim predlažemo da se naprave dopunska istraživanja kojima bi se utvrdilo je li potrebno uvesti novu klasu na trećoj razini NKS-a ili redefinirati postojeću klasu ili ime klase na trećoj razini.
Biševo
Otok Biševo je jedan od biljnogeografski najzanimljivijih otoka. U formiranju trenutnih staništa na otoku Biševu veliku ulogu je imao požar 2003. godine, koji je zahvatio više od 3/4 otoka. Samo krajnji sjeverni dio otoka nije izgorio. Stoga trenutno na otoku Biševu dominiraju „bušici ružmarina s mnogocvjetnom resikom“ (Erico-Rosmarinetum H-ić. 1958). Tako su neke površine više, a neke manje zahvaćene ponovnom sukcesijom bušika. Na površinama na kojima još nema toliko ružmarina i resike nalazimo elemente „eu- i stenomediteranskih pašnjaka raščice“, koji su uglavnom razvijeni u obliku monodominantnosti vrste Brachypodium retusum. Na područjima na kojima vegetacija nije potpuno izgorjela, nalazimo manje površine drugih stanišnih tipova, ali i oni uglavnom dolaze s većim ili manjim udjelom bušika. Na krajnjem sjeveru otoka i u središnjem dijelu nalazi se „čista, vazdazelena šuma i makija crnike s mirtom“ (Myrto-Quercetum ilicis (H-ić)Trinajstić 1985). U sjevernom i središnjem dijelu otoka postoje manje površine na kojima je sačuvana šuma alepskog bora. Ovu šumu nalazimo u 2 stanišna oblika – kao „šumu alepskog bora sa sominom“ (Junipero phoeniceae-Pinetum halepensis Trinajstić 1988) i „šumu alepskog bora s tršljom“ (Pistacio-Pinetum halepensis De Marco, Veri et Caneva 1984). U zapadnom dijelu otoka se uz obalu na nekoliko mjesta nalaze elementi “makije tršlje i somine” (Pistacio-Juniperetum phoeniceae Trinajstić 1987) miješani s bušikom. Na stijenama koje se nalaze na južnoj strani otoka razvila se vegetacija “dalmatinskih vapnenačkih stijena” (Centaureo-Portenschlagiellion Trinajstić 1980). U središnjem dijelu, kod Biševog polja i Potoka, te na sjeveru kod Sablunare, postoje još vinogradi koji se obrađuju. Uglavnom se radi o malim vinogradima između kojih su napuštena polja obrasla bušikom. Ostale, nekoć obradive površine, uglavnom su napuštene, te obraštaju bušikom, ruderalnom vegetacijom i kupinom.
Svetac (Sveti Andrija)
Otok Svetac je najšumovitiji i najzeleniji otok Viškog arhipelaga. Zbog aridnije klime od otoka Visa i Biševa, na Svecu nalazimo veće površine pod stenomediteranskom þistom vazdazelenom šumom i makijom crnike (Oleo-Ceratonion). Najveći dio otoka Svetca prekrivaju šume alepskog bora (Pinus halepensis). U najvišem dijelu otoka nalazi se „mješovita šuma alepskog bora i crnike“ (Querco ilicisPinetum halepensis Loisel 1971). Na nekoliko mjesta je na južnim padinama razvijena „šuma alepskog bora s tršljom“ (Pistacio-Pinetum halapensis De Marco, Veri et Caneva 1984), u kojoj je Pistacia lentiscus obilno razvijena u sloju grmlja. Ostale šume alepskog bora, koje se uglavnom nalaze na sjevernoj strani otoka, opisane su kao stanišni tip „šuma alepskog bora sa sominom“ (Junipero phoeniceae-Pinetum halapensis Trinajstić 1988), zbog većeg udjela crnike u sloju grmlja. Na južnoj strani otoka Sveca prevladava makija u kojoj dominiraju somina (Juniperus phoenicea) i tršlja (Pistacia lentiscus Br.-Bl. 1931), koja je na ovoj karti opisana kao „makija divlje masline i somine“ (Oleo-Juniperetum phoenicaea Bruno et. al. 1983), premda u nije nađena divlja maslina.
Ovaj tip makije često mozaično raste s „bušikom ružmarina s mnogocvjetnom resikom“ (EricoRosmarinetum H-ić 1958). Premda nekoć naseljen otok, danas ima samo nekoliko malih vinograda koji se još obrađuju. Površine koje su se prije obrađivale, većinom terase s vinogradima, danas su uglavnom zarasle u bušik i makiju. U priobalnom dijelu zone zračne posolice i prskanja morskih valova prevladava stanišni tip „površine stjenovitih obala pod halofitima“ (Critmo-Limonion Br.-Bl. Molinier 1934). Na strmim, okomitim stijenama uz morsku obalu prevladava hazmofistka vegetacija stijenjača pukotinjarki „dalmatinskih vapnenačkih stijena“ (Centaureo-Portenschlagiellion Trinajstić 1980). U sjevernom dijelu otoka na nekoliko mjesta je nađena i Euphorbia dendroides, te su te stijene opisane kao mozaik „dalmatinskih vapnenačkih stijena“ i „ljeti listopadnih grmljaka“.